Какие пигменты содержатся в хлоропластах и какова их роль

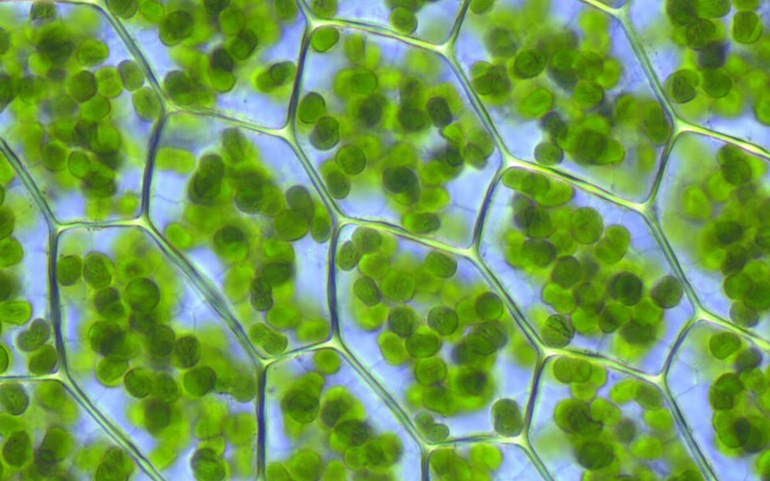
Хлоропла́сты (от греч. χλωρός — «зелёный» и от πλαστός — вылепленный) — зелёные пластиды, которые встречаются в клетках фотосинтезирующих эукариот. С их помощью происходит фотосинтез. Хлоропласты содержат хлорофилл. У зелёных растений являются двумембранными органеллами[Пр. 1]. Под двойной мембраной имеются тилакоиды (мембранные образования, в которых находится электронтранспортная цепь хлоропластов). Тилакоиды высших растений группируются в граны, которые представляют собой стопки сплюснутых и тесно прижатых друг к другу тилакоидов, имеющих форму дисков. Соединяются граны с помощью ламелл. Пространство между оболочкой хлоропласта и тилакоидами называется стромой. В строме содержатся хлоропластные молекулы РНК, пластидная ДНК, рибосомы, крахмальные зёрна, а также ферменты цикла Кальвина[1].
Происхождение[править | править код]
В настоящее время общепризнано[2] происхождение хлоропластов путём симбиогенеза.
Предполагают, что хлоропласты возникли из цианобактерий, так как являются двумембранным органоидом, имеют собственную замкнутую кольцевую ДНК и РНК, полноценный аппарат синтеза белка (причем рибосомы прокариотического типа–70S), размножаются бинарным делением, а мембраны тилакоидов похожи на мембраны прокариот (наличием кислых липидов) и напоминают соответствующие органеллы у цианобактерий. У глаукофитовых водорослей вместо типичных хлоропластов в клетках содержатся цианеллы — цианобактерии, потерявшие в результате эндосимбиоза способность к самостоятельному существованию, но отчасти сохранившие цианобактериальную клеточную стенку[3].
Давность этого события оценивают в 1 — 1,5 млрд лет[4].
Часть групп организмов получала хлоропласты в результате эндосимбиоза не с прокариотными клетками, а с другими эукариотами, уже имеющими хлоропласты[5]. Этим объясняется наличие в оболочке хлоропластов некоторых организмов более чем двух мембран[Пр. 2]. Самая внутренняя из этих мембран трактуется как потерявшая клеточную стенку оболочка цианобактерии, внешняя — как стенка симбионтофорной вакуоли хозяина. Промежуточные мембраны — принадлежат вошедшему в симбиоз редуцированному эукариотному организму. У некоторых[Пр. 3] групп в перипластидном пространстве между второй и третьей мембраной располагается нуклеоморф, сильно редуцированное эукариотное ядро[6].
Строение[править | править код]
1. наружная мембрана
2. межмембранное пространство
3. внутренняя мембрана (1+2+3: оболочка)
4. строма (жидкость)
5. тилакоид с просветом (люменом) внутри
6. мембрана тилакоида
7. грана (стопка тилакоидов)
8. тилакоид (ламела)
9. зерно крахмала
10. рибосома
11. пластидная ДНК
12. пластоглобула (капля жира)
У различных групп организмов хлоропласты значительно различаются по размерам, строению и количеству в клетке. Особенности строения хлоропластов имеют большое таксономическое значение[7]. В основном хлоропласты имеют форму двояковыпуклой линзы, размер их около 4-6 мкм.
Оболочка хлоропластов[править | править код]
У различных групп организмов оболочка хлоропластов отличается по строению.
У глаукоцистофитовых, красных, зелёных водорослей[8] и у высших растений оболочка состоит из двух мембран. У других эукариотных водорослей хлоропласт дополнительно окружён одной или двумя мембранами. У водорослей, обладающих четырёхмембранными хлоропластами, наружная мембрана обычно переходит в наружную мембрану ядра.
Перипластидное пространство[править | править код]
Граны[править | править код]
Граны представляют собой стопки дисковидных тилакоидов. Отдельные граны хлоропласта соединятся более длинными ламеллами, которые также называют межграновыми или строматическими тилакоидами. Грановые и межграновые тилакоиды различаются белковым составом.
Пиреноиды[править | править код]
Пиреноиды — центры синтеза полисахаридов в хлоропластах[9]. Строение пиреноидов разнообразно, и не всегда они морфологически выражены. Могут быть внутрипластидными и стебельчатыми, выступающими в цитоплазму. У зелёных водорослей и растений пиреноиды располагаются внутри хлоропласта, что связано с внутрипластидным запасанием крахмала.
Стигма[править | править код]
Стигмы, или глазки, встречаются в хлоропластах подвижных клеток водорослей. Стигмы содержат каротиноиды и состоят из липидных глобул. Располагаются вблизи основания жгутика и вместе с особым вздутием на нём выполняют роль фоторецептора, задействованного в осуществлении клеточного фототаксиса[10].
См. также[править | править код]
- Фотосинтез
- Триозофосфатный транслокатор
- Хромопласты
- Цианеллы
Примечания[править | править код]
Комментарии[править | править код]
- ↑ Хлоропласты организмов, относящихся к группе хромистов, имеют четырёхслойную оболочку. Предполагается, что в истории их возникновения включение одной клетки в состав другой происходило дважды.
- ↑ Например, у динофитовых и эвгленовых имеется 3 мембраны, а у охрофитов — 4.
- ↑ У криптофитовых, хлорарахниофитовых и некоторых динофитовых.
Примечания[править | править код]
- ↑ Тихонов А. Н. Трансформация энергии в хлоропластах — энергопреобразующих органеллах растительной клетки // Соровский Образовательный Журнал. 1996. № 4. С. 24—32
- ↑ Карпов, 2001, с. 246.
- ↑ Карпов, 2001, с. 249,246.
- ↑ Белякова, 2006, с. 35.
- ↑ Карпов, 2001, с. 249.
- ↑ Карпов, 2001, с. 250.
- ↑ Карпов, 2001, с. 235.
- ↑ Белякова, 2006, с. 32—34.
- ↑ Карпов, 2001, с. 239.
- ↑ Карпов, 2001, с. 240.
Литература[править | править код]
- Белякова Г. А. Водоросли и грибы // Ботаника: в 4 т. / Белякова Г. А., Дьяков Ю. Т., Тарасов К. Л. — М.: Издательский центр «Академия», 2006. — Т. 1. — 320 с. — 3000 экз. — ISBN 5-7695-2731-5.
- Карпов С.А. Строение клетки протистов. — СПб.: ТЕССА, 2001. — 384 с. — 1000 экз. — ISBN 5-94086-010-9.
- Lee, R. E. Phycology, 4th edition. — Cambridge: Cambridge University Press, 2008. — 547 с. — ISBN 9780521682770.
- ХЛОРОПЛАСТЫ // Большая российская энциклопедия. Электронная версия (2017); https://bigenc.ru/biology/text/4694635 Дата обращения: 23.06.2018
Функциональные особенности
Строение хлоропласта изучается школьниками в 6 классе на уроках биологии. К особенностям клеток относится наличие в строме рибосомы, ДНК, РНК. В мембране присутствует вещество, способное придать растениям соответствующий цвет. Для хлорофилла характерен зеленый оттенок, а для каротиноида:
- красный;
- желтый;
- оранжевый.

Значение хлорофилла для растений заключается в возможности осуществления процесса фотосинтеза. С учётом строения биологи выделяют 4 типа хлорофилла: a, b, c, d. Первые два содержатся в растениях на суше и зеленых водорослях. Типы a и c считаются растительными компонентами диатомовых, d и a — красных водорослей.
Для хлорофилла характерно поглощение солнечной энергии с последующей передачей иным молекулам. Разрушение зеленого вещества наблюдается в конце жизненного цикла органоида в результате резкого изменения светового дня и значения температуры. Часть хлоропластов превращается в хромопласты. Это приводит к изменению внутренней информации, появлению нового цветового оттенка, опадению листьев.
Принципы классификации
Пластиды делятся на три вида: лейкопласты (бесцветные), хлоропласты (окрашенные в зеленый цвет), хромопласты (имеют разные оттенки). На протяжении жизни клетки способны превращаться друг в друга. Лейкопластам свойственно переходить в хлоропласты, а последние за счёт появления бурых и прочих пигментов — в хромопласты, пластоглобулы.
Внешне зеленые вещества покрыты липидной и белковой мембранами. Полужидкая строма с тилакоидами (компартменты, ограниченные мембраной) считается основным веществом, в состав которого входят граны с каналами. Первые компоненты представлены в виде плоских круглых мешочков, расположенных перпендикулярно поверхности двухмембранных органоидов (ДО).
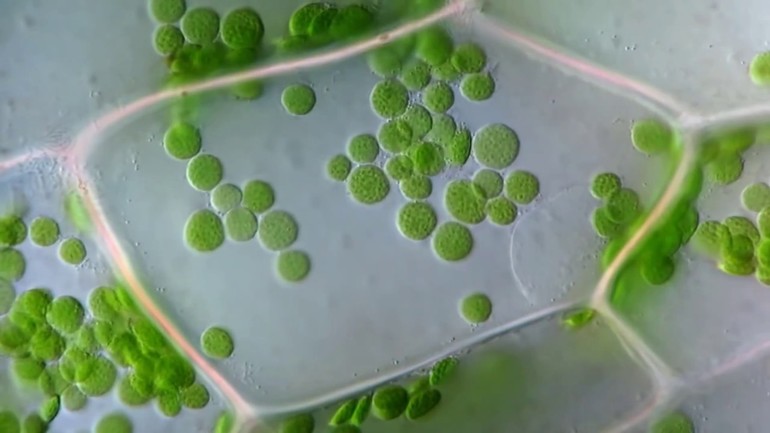
Уникальность их структуры заключается в хранении зеленого пигмента (хлорофилл). Главная функция хлоропластов связана с участием в фотосинтетическом явлении. В их состав входят жиры, зерна (митохондрия, пропластида), крахмал.
На долю липидов приходится до 30%. Они представлены тремя группами:
- Структурная. В состав входят амфипатические вещества.
- Гидрофобная. В группу входят каротиноиды, которые защищают зеленые вещества от фотоокисления. Одновременно они транспортируют водород.
- Жирорастворимая. Группа состоит из витаминов К и Е.
К другим компонентам, входящим в состав хлоропласта, относятся углеводы. Они представлены в виде продуктов фотосинтеза. До 25% приходится на долю минералов. Ферменты могут выполнять двойную функцию: катализацию различных реакций, обеспечение биосинтеза белков.
Внутренняя структурированность хлоропластов зависит от функциональных нагрузок, физиологического состояния. Молодые клетки размножаются за счет деления, а зрелые обладают выраженной системой гран. Если они стареют, происходит разрыв тилакоидов, распадается хлорофилл. Осенью деградация приводит к появлению хромопластов.
Главная роль хлоропластов в фотосинтезе обеспечена их способностью пассивно двигаться в клетках, увлекаемых током цитоплазмы. Веществу свойственно собирать свет и активно перемещаться с одного места на другое. При интенсивном свете оно поворачивается ребром к яркому солнцу, выстраиваясь вдоль стенок, которые параллельны лучам.
Если освещение слабое, схема движения хлоропластов следующая: они перемещаются на стенки, обращённые к солнцу, поворачиваясь наибольшей поверхностью. Когда освещение среднее, клетки занимают соответствующее положение. От условий освещения зависит то, какие пигменты хлоропластов появятся.
Для пластид и митохондрий свойственна полуавтономная степень. Кроме фотосинтеза, в первых компонентах происходит биосинтез белка. Так как они содержат в себе ДНК, поэтому принимают активное участие в наследственном комплексе: передача признаков, цитоплазматические свойства.
Описание хромопластов
К пластидам высших растений относятся хромопласты. Они имеют незначительные размеры. Для внутриклеточных органелл характерен разный окрас: красный, желтый, коричневый. Он придает соответствующий цвет осенью, плодам и цветкам, что необходимо для привлечения опылителей и животных, разносящих семена продолжительные расстояния.
Структура ткани похожа на иные пластиды. Внутренняя оболочка развита слабее внешней. У некоторых представителей она может отсутствовать. В каротиноидах (жирорастворимые пигменты) происходит накапливание кристаллов. Для определения точных функций вещества изучается таблица с формами хромопластов:
- многоугольная;
- овальная;
- серповидная;
- игольчатая.

Их роль в жизни растений до конца не выяснена. Ученые предполагают, что пигменты участвуют в окислительных и восстановительных процессах, необходимых для размножения и физиологического развития клеток.
Строение лейкопластов
В органоидах этого типа накапливаются питательные компоненты. Лейкопласты имеют 2 оболочки: внутреннюю и внешнюю. На свету им свойственно превращаться в хлоропласты, но в привычном состоянии органоиды бесцветны. Основная их форма — шаровидная. Размещены они в мягких частях растений:
- стебель;
- корень;
- луковица;
- листья.
С учетом накапливаемого вещества лейкопласты классифицируются на следующие виды: амилопласты, элайопласты, протеинопласты. В первую группу входят органоиды с крахмалом, находящиеся в каждом растении. Если лейкопласт полностью заполнен крахмалом, он называется крахмальным зерном. Для элайопластов характерно продуцирование и запас жиров, а для протеинопластов — скопление белковых веществ.
Лейкопласты обладают ферментной субстанцией, что способствует ускоренному протеканию химических реакций. В отрицательном жизненном периоде, когда не происходит фотосинтез, они расщепляют полисахариды на простые углеводы. Так как в луковицах содержится много органоидов, поэтому им свойственно переносить длительную засуху, жару, низкую температуру. После выполнения своих функций они становятся хромопластами.
Симбиотическая теория
Чтобы выяснить механизм появления пластид, митохондрий и других органоидов, рассматривается теория эндосимбиоза. Ее суть заключается в совместной и взаимовыгодной жизни органеллы с клеткой. Впервые теорию предложил Шимпер в 1883 году. В 1867 ученые работали над двойственной природой лишайников.
Биолог Фамицын, учитывая теорию Шимпера, предположил, что хлоропласты, как лишайники и водоросли, относятся к симбионтам. Ученые доказали, что митохондрии — аэробные бактерии, которые не размножаются за пределами клеток. Общие свойства, характерные для митохондрий и пластид:
- наличие двух замкнутых мембран;
- размножение бинарным делением;
- ДНК не связана с гистонами;
- наличие своего аппарата синтеза белка.
В ДНК пластид и митохондрий, в отличие от аналогичных структур прокариот, нет интронов. А в ДНК хлоропластов закодирована информация о некоторых белках, остальные данные находятся в ядре клетки. В результате эволюции часть генетического материала из генома перешло в ядро, поэтому хлоропласты и митохондрии не размножаются независимо.
Археи и бактерии не склонны к фагоцитозу. Они питаются только осмотрофно. Множественные биологические и химические исследования указывают на химерную сущность бактерий. Ученые не выяснили, как сливаются организмы из нескольких доменов. В условиях современности выявлены организмы, которые содержат в себе другие клетки в качестве эндосимбионтов. Они отличаются от первичных эукариотов тем, что не интегрируются в одно целое, не имеют своей индивидуальности.
Интересным организмом считается Mixotricha paradoxa. Чтобы двигаться, она использует 250 000 бактерий, которые фиксируются на ее поверхности. Митохондрии у этого организма вторично потеряны. Внутри находятся сферические аэробные микроорганизмы, которые заменяют органеллы.
Фотосинтез протекает при помощи пигментов зеленых листьев. Последние содержат два хлорофилла — а (голубовато-зеленый) и б (желтовато-зеленый) (см. главу Пиррол ) — и две группы каротиноидов — каротины (оранжевые) и ксантофиллы (желтые). Эта система пигментов находится в хлоропластах — частицах удлиненной формы, находящихся в клетках зеленых листьев. Установлено, что лучистая энергия, поглощенная одним пигментом, может быть передана другому пигменту хлоропласта. При облучении хлоропластов светом с длиной волны, поглощаемой исключительно хлорофиллом б, испускаемое излучение (за счет флуоресценции) содержит длины волн, характерные для хлорофилла а, в то время как флуоресценция хлорофилла б уменьшается. Остальные пигменты клетки могут передавать аналогичным образом поглощенную энергию хлорофиллу а. Тем самым расширяется спектральная область, потребляемая в фотосинтезе. Хлорофилл передает поглощенную лучистую энергию химической системе при помощи еще не выясненного механизма. [c.260]
Пигменты хлоропластов относятся к двум основным группам — хлорофиллам и каротиноидам (каротины и ксантофиллы). В нормальных зеленых листьях хлорофилла много больше, чем каротиноидов их соотношение иногда достигает 5 1. [c.257]
Отношение оптических плотностей в зеленом минимуме и красном максимуме кривых поглощения составляет (см. табл. 2) меньше чем 0,01 в чистом хлорофилле а и около 0,05 в эфирном экстракте из листьев ячменя, содержащем все пигменты хлоропластов однако, как видно из табл. 19, это отношение в живых водорослях превышает 0,3,. а в зеленых листьях достигает 0,6. [c.116]
Опыты, выполненные М. С. Цветом, позволили ему не только разделить смесь растительных пигментов (хлоропластов) на отдельные компоненты, но и сформулировать ряд важных теоретических положений, объясняющих причины разделения смесей некоторых веществ при помощи хроматографии. М. С. Цвет показал, что вещества, растворенные в определенной жидкости, при прохождении через колонку, заполненную адсорбентом, образуют адсорбционный ряд, выражающий относительное адсорбционное сродство членов этого ряда к адсорбенту. Согласно закону адсорбционного замещения, сформулированному М. С. Цветом, каждый член адсорбционного ряда, обладающий большим адсорбционным сродством, чем последующий, вытесняет его из адсорбента и, в свою очередь, вытесняется предыдущим. Таким образом, распределение веществ в колонке на зоны выражает относительное положение этих соединений в адсорбционном ряду. [c.31]
Проведенные после Блэкмана многочисленные наблюдения показывают, что при благоприятных условиях температурный коэффициент (Сю) процесса фотосинтеза достигает нередко 2 и выше 2. Существование в цепи процессов фотосинтеза чисто химических звеньев и является причиной того, что низкие температуры снижают фотосинтетическую активность растения даже в условиях интенсивного освещения. Крайне неустойчив фотосинтез и по отношению к веществам, ингибирующим дыхание (эфир, фенилуретан, цианиды), причем полное подавление фотосинтеза имеет место при применении ядов в таких концентрациях, которые не только не оказывают влияния на пигменты хлоропластов, но оставляют нетронутым и дыхание тех же листьев. [c.141]
Направляя на зеленый лист монохроматический свет различной длины волны, полученный с помощью светофильтра, призмы или дифракционной решетки, и измеряя скорость фотосинтеза, соответствующую каждой длине волны, можно убедиться в том, что свет в синей ( 420 нм) и красной ( 670 нм) областях спектра обеспечивает наибольшую эффективность фотосинтеза, а свет в зеленой 500—600 нм) области—г наименьшую (рис. 4.7). Такой спектр действия (зависимость относительной эффективности процесса от длины волны) можно объяснить, исходя из спектра поглощения хлорофилла, главного пигмента хлоропластов. Экстрагированный из листьев хлорофилл интен- [c.114]
Пигменты хлоропластов. В процессе фотосинтеза происходит поглощение света пигментами, сосредоточенными в хлоропластах. Пигменты зеленых растений играют важную роль в превращении лучистой энергии света в химическую. У авто-трофных растений уменьшение количества пигментов приводит [c.157]
Участие пигментов хлоропластов в процессах жизнедеятельности растения [c.159]
В процессе фотосинтеза происходит поглощение световой энергии пигментами хлоропласта и ее превращение в химическую. Поглощенная энергия передается на реакционные центры. Минимальное число молекул хлорофилла с вспомогательными пигментами, обслуживающими реакционный центр, называется фотосинтетической единицей. Такая единица содержит у растений 250—400 молекул хлорофилла, вмонтированных в белково-липоидную мембрану. [c.127]
Большая часть белков листьев локализована в хлоропластах и представляет собой белок фракции I, который участвует в фиксации СО2 (цикл Кальвина). Пигменты хлоропластов представляют собой хромопротеиды, а многие другие белки, специфичные для хлоропластов, являются ферментами, которые катализируют реакции, протекающие только в хлоропластах. Белки (главным образом ферменты) присутствуют и в других органеллах и в цитоплазме клеток листа. [c.44]
Разделение и хроматография пигментов хлоропластов [c.122]
Циклические каротиноиды могут иметь эпоксигруппы при С-5,6, например в пигменте хлоропластов, виолаксантине [5,6, 5, 6 -диэпокси-5,6, 5,6 -тетрагидро-р,р-каротин-3,3 -диол (2.8)]. [c.38]
Пигменты хлоропластов служат исключительно переносчиками энергии (фотохимические сенсибилизаторы) для осуществления эндэргонного разложения воды. [c.261]
Хлоропласты у разных растений имеют самую разнообразную форму, но, как правило, они гораздо крупнее митохондрий (рис. 23-6). Снаружи хлоропласты окружены непрерывной довольно хрупкой наружной мембраной. Внутренняя мембранная система ограничивает собой внутренний компартмент хлоропласта. В нем находится много плоских мембранных мешочков, или пузырьков, часто связанных с внутренней мембраной. Это так называемые тилакоиды они обычно собраны в стопки, называемые гранами (рис. 23-6). В тилакоидных мембранах содержатся все фотосинтетиче-ские пигменты хлоропласта и все ферменты, необходимые для первичных световых реакций. Большинство ферментов, участвующих в темновых реакциях, в которых СОа восстанавливается до глюкозы, находится в жидкости, заполняющей внутренний компартмент хлоропласта и окружающей тилакоиды ее называют стромой. У многих видов темновые реакции протекают также и [c.687]
A. Н. Теренина, А. А. Ерасновского, Т. Н. Годнева, А. А. Табенц-кого, Н. М. Сисакяна, О. П. Осиповой дали материалы решающего значения для познания природы пигментов хлоропластов, их оптических свойств, состояния в растениях, механизма участия в процессе фотосинтеза, химизма образования хлорофилла . [c.12]
Когда листья помещают в эфир, или охлаждают жидким воздухом,, или кипятят в воде, полоса поглощения смещается к положению, соответствующему истинному раствору это указывает на вероятное разложение белково-хлорофильного комплекса (Вильштеттер и Штоль [119] Зейбольд и Эгле [156]). В убитых таким способом листьях хлорофилл гораздо чувствительнее к кислороду и кислотам, чем до убивания. Таким образом, представ-вдется вероятным, что хлорофилл и другие пигменты хлоропластов связаны в живой клетке с клеточными белками, а также с некоторыми липофильными соединениями. Возникает вопрос, осуществляется ли эта связь в стехио-метрическом отношении и затрагивает ли она в равной мере весь хлорофилл, содержащийся в клетке [c.386]
Если определено полное поглощение света в листе, слоевище или суспензии клеток, то возникает вопрос, какая же часть этого поглощения приходится на долю пигментов, находящихся в хлоропластах. Многие авторы (начиная с Рейнке в 1886 г. [6]) допускали, что определенная часть поглощения белого света в растениях приходится на долю бесцветных частей тканей — цитоплазмы, клеточного сока, зерен крахмала и целлюлозы. Зейбольд произвольно отнес /g часть полного поглощения на долю поглощения этих компонентов и % приписал пигментам хлоропластов. Кривая поглощения бесцветного листа герани, данная Зейбольдом и Вейссвейлером [43], показывает значительное поглощение вблизи сине-фиолетового конца видимого спектра. Несомненно, что истинно бесцветные вещества не могут поглощать видимый свет. Однако растительные клетки содержат окрашенные вещества, связанные с оболочками клеток или с клеточным соком, а не с пластидами к ним относятся флавоны, таннины и т. п. Некоторые из этих веществ слабо окрашены и обычно имеют желтый цвет, другие, хотя и имеют интенсивную окраску, присутствуют в очень малых концентрациях по сравнению с пигментами пластид. У некоторых видов, однако, флавоны и антоцианины присутствуют в таком количестве, что придают листьям яркокрасный цвет (листья красных разновидностей и молодые листья многих растений весаой). Цвет этих листьев свидетельствует о том, что значительная часть поглощенной ими световой энергии приходится на долю непластидных пигментов. [c.92]
Необходимо учитывать, что усвоение углекислого газа является лишь одним из процессов, в ходе которых происходит запасание энергии света, поглощенного пигментами хлоропластов. Прямим доказательством этого является зарегистрированное с помощью высокочувствительных фотомикрокалориметров нарушение между величиной запасаемой энерши и интенсивностью выделения Ог в процессе фотосинтеза у хлореллы, вызванное изменением спектрального состава света (Белл, 1965) или температуры (Петров, Кайбияйнен, 1%9). [c.256]
В квантосомах ликаишзованы все пигменты хлоропластов. Соотношение пигментов и хинонов примерно такое же, как в хлоро- [c.103]
Пигменты хлоропластов хлорофиллы, цитохромы и каротиноиды в природном состоянии, очевидно, не являются свободными, но связаны с протеинами и липопротеинами. Фрей-Вислинг и Штейнман (1957) установили, что па хороших электронных микрофотографиях ламеллы могут рассматриваться как состоящие из сферических частиц около 65 А в диаметре и что эти частицы представляют собой макромолекулы протеинов и [c.308]
Установленная в данной работе малая инер-ционность процессов высвобождения и исчезновения подвижных электронных вакансий (положительных дырок) в пленках хлорофилла, хлорофиллида и пигментов хлоропластов показывает, что мы имеем дело с первичным процессом. После первоначального захвата электрона от возбужденной молекулы ловушками, имеющимися даже в ненарушенном кристалле пигмента, происходит эстафетный обмен электроном между нижними — основными (синглетными) уровнями молекул [И, 27, 29]. В этой картине явления мы следуем представлениям, развитым Лайонсом для дырочной фотопроводимости кристаллов антрацена [30]. [c.280]
Высоконенасыщенные тетратерпены, обнаруженные в высших растениях, грибах, фотосинтезирующих бактериях и водорослях, обычно входят в состав сложных смесей фотосинтезирующих лигментов. Эти так называемые Пигменты хлоропластов представляют собой интенсивно окрашенные соединения, что и послужило главной причиной их выбора в качестве объектов на которых впервые был продемонстрирован принцип хроматографического разделения Фракционирование терпенов этой группы проводилось на самых разнообразных сорбентах, в том числе и таких достаточно распространенных, как силикагель, оксид алюминия, карбонаты цинка и кальция, оксид магния, гидроксид кальция, целлюлоза и сахароза. Элюентами обычно слуЯсили смеси углеводородов, содержащие более полярный растворитель, -например диэтиловый эфир, ацетон, метанол или пропанол. [c.248]
Большие успехи достигнуты в области использования ВЭЖХ для разделения пигментов хлоропластов и других растительных пигментов (см. также ч. 2, гл. 10). Благодаря высокой скорости эксперимента и хорошей воспроизводимости результатов ВЭЖХ почти вытеснила другие методы хроматографии. При разделении на колонках с мелкодисперсным силикагелем [353 элюентом обычно служит гексан, содержащий пропанол-1 [354 или пропанол-2 [355]. С помощью хроматографии на колонке с ц-партисилом 10 в системе легкий петролейный эфир— аце-тон — диметилсульфоксид — диэтиламин (300 93 6 1) можно [c.248]
Хайфлосуперцел нашел применение и в тонкослойной хроматографии. Смесь этого сорбента с оксидом магния и сульфатом кальция образует прочные слои, элюирование на которых можно проводить с высокой скоростью. Разделение неполярных каротинов на таких слоях протекает с хорошим разрешением [369]. На таких слоях можно, например, за один прием разделить все каротиноиды моркови. Хроматографический анализ пигментов хлоропластов перца apsi um включает две стадии сначала на пластинках с целлюлозой разделяют хлорофиллы и ксантофиллы, а затем смесь каротинов хроматографируют на тонком слое оксида магния и хайфлосуперцела [370]. На слоях из оксида магния элюирование можно вести легким петролейным эфиром. ТСХ на этом сорбенте в сочетании с УФ-детектиро-ванием составляет основу метода количественного определения а- и р-каротинов в биомассе [371]. Для обнаружения небольших количеств р-каротина предложен метод хроматографии в тонком слое, сформированном из смеси карбоната кальция, оксида магния и гидроксида кальция, в системе ацетон — легкий петролейный эфир — хлороформ (5 5 4) [372]. Проведен сравнительный анализ эффективности разделения главных пигментов на слоях из кукурузного крахмала, целлюлозы и микрокристаллической целлюлозы. В системе гептан — этилацетат — пропанол полное разделение было достигнуто на слоях из крахмала [373]. [c.251]
Растительные пигменты хроматографируют также в тонком слое силикагеля [374], однако этот сорбент, по-видимому, более подходит для выделения всей группы пигментов хлоропластов, чем для их разделения на индивидуальные компоненты. Так, например, на силикагеле, содержаш,ем 10% сульфата аммония, можно лишь отделить каротины и феофитины от хлорофиллов, а разделить каротины практически не удается. Используя высокоскоростной видеоденситометр, аналогичный применяемому в тех аминокислот, можно оценивать количество вещества в каждом хроматографическом пятне, причем результаты такого анализа близки к полученным обычными спектрометрическими методами. Проводя элюирование смесями грет-бутанол — бензол (1 9) и грег-бутанол — пентан — ацетон (1 18 1), авторам работы [377] удалось быстро и достаточно легко отделить хлорофиллы и их производные от желтых каротиноидных пигментов. В системе дихлорметан — этилацетат — диэтиловый эфир (8 2 1) была полностью разделена смесь зеаксантина, лютеина и диэфира лютеина [377]. Индивидуальные компоненты элюировали с силикагеля этанолом и определяли их фотометрически. Хроматографическая подвижность астаксантина и кантак- [c.251]
Интересной модификацией метода ТСХ является обращенно-фазовая ТСХ [379]. На химически связанной фазе is (ODS) в системе метанол — ацетон — вода (20 4 3) была разделена смесь пигментов хлоропластов шпината, содержащая неоксантин, хлорофиллы аиЬ, каротин, виолаксантин и лютеины. Этим же методом были разделены каротиноиды цитрусовых [380]. [c.253]
Согласно современным представлениям, в тилакоид-ных мембранах локализованы все фотосиитетические пигменты хлоропласта и ферменты, необходимые для осуществления световых реакций фотосинтеза. В строме содержатся ферменты, участвующие в темновых превращениях диоксида углерода. Таким образом, сложная и тонкая структура хлоропласта обеспечивает пространственное разделение отдельных реакций а тем самым и эффективный ход фотосинтеза в целом. Образующиеся в пластидах продукты ассимиляции траиспор- [c.74]
При обработке мембран тилакоидов детергентами можно перевести в раствор те белки, которые обычно тесно связаны с мембранами. Растворенные таким образом компоненты можно разделить по их молекулярным массам, используя, например, электрофорез в полиакриламидном геле. Полосы, соответствующие отдельным белковым компонентам, проявятся после обработки геля специальными красителями. Наряду с этим можно получать мутантов растений и водорослей, не имеющих специфических белков или пигментов хлоропластов и вместе с тем утративших какие-либо характерные функции или фотосинтетическую активность в целом. Сравнивая набор компонентов, обнаруживаемых в геле после электрофореза белков из мутантов и из организмов дикого типа, можно установить зависимость между определенной функцией и тем или иным компонентом, например белком или пигмент-белковым комплексом. Таким образом были разделены и предварительно охарактеризованы связанный с реакционным центром фотосистемы I комплекс Руоо—хлорофилл а—белок связанный с реакционным центром фотосистемы II комплекс светособирающий хлорофилл а/Ь — белок железо-серные белки, связанные с мембраной цитохром / АТРаза, или сопрягающий фактор, и другие компоненты мембран тилакоидов (см. рис. 8. 1). [c.66]