Какие нейроны не содержатся в ганглиях интрамуральныйх сплетений

Материал взят с сайта www.hystology.ru
Вегетативный отдел нервной системы включает высшие вегетативные центры, локализованные в промежуточном мозге в области III желудочка, вегетативные ядра серого вещества ствола и спинного мозга, а также периферические нервные ганглии.
В отличие от рефлекторных дуг соматической нервной системы моторный нейрон рефлекторных дуг вегетативного отдела идет не в составе серого вещества центральной нервной системы, а в одном из периферических ганглиев.
Вегетативная нервная система представлена парасимпатической и симпатической системами. К парасимпатической нервной системе относят нейроны черепно-мозгового и крестцового участков мозга и связанные с ними ганглии. Симпатическая нервная система объединяет нейроны грудопоясничного отдела мозга и связанные с ними превертебральные и паравертебральные ганглии. Симпатические нервные волокна иннервируют все органы, тогда как парасимпатическая нервная система иннервирует лишь органы, развившиеся из эмбриональной кишки или в связи с пей.
Большинство внутренних органов получает как симпатические, так и парасимпатические нервные волокна.
Эфферентное звено вегетативных рефлекторных дуг двухчленно. Центральный нейрон всегда располагается в головном или спинном мозге. Второй нейрон (периферический) в симпатической нервной системе лежит в превертебральных или паравертебральных узлах, а в парасимпатической — в органе или вблизи него (рис. 180).
В парасимпатической нервной системе оба нейрона холинергические. Пресинаптический полюс как первого, так и второго нейрона содержит мелкие, светлые синаптические пузырьки (40 — 60 нм) и одиночные крупные электроноплотные везикулы.
В симпатической нервной системе первый нейрон холинергический, второй адренергический (медиатор — норадреналин). Морфологически адренергические синаптические пузырьки (30 — 50 нм в диаметре) характеризуются электроноплотной сердцевиной (гранулярные синаптические пузырьки). Кроме характерных для адренергических синапсов гранулярных везикул, в их составе, как и в холинергических, выявляют и большие гранулярные
Рис. 180. Простая вегетативная рефлекторная дуга:
1 — спинной мозг; 2 — спинномозговой узел; 3 — передний корешок; 4 — задний рог; 5 — боковой рог; 6 — передний рог; 7 — чувствительный (афферентный) нейрон соматической и симпатической нервной системы; 8 — центральный (эфферентный) нейрон вегетативной нервной системы; 9 — двигательный (эфферентный) нейрон передних рогов; 10 — узел симпатического ствола; 11 — узел солнечного сплетения; 12 — интрамуральный узел (узел нервного сплетения пищевода); 13, 14, 15 — периферические (эфферентные) нейроны вегетативной нервной системы; 16 — преганглионарные волокна эфферентного пути вегетативной нервной системы; 17 — постганглионарные волокна эфферентного пути вегетативной нервной системы; 18 — эфферентный путь соматической нервной системы; 19 — стенка пищевода; 20 — поперечнополосатые мышцы; 21 — микроскопическое строение периферических узлов вегетативной нервной системы; 22 — мультиполярная нервная клетка; 23 — клетки глии; 24 — нервное волокно.

Рис. 181. Нервные клетки вегетативного ганглия:
1 — длинноаксонная нервная клетка; 2 — аксон; 3 — равноотростчатая нервная клетка; 4 — ядра клеток глии.
везикулы диаметром 60 — 120 им (1 — 5 % к общему количеству).
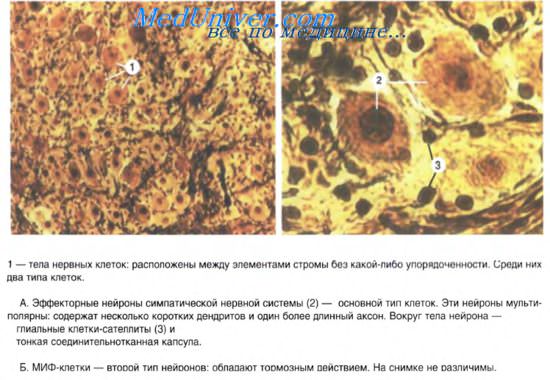
В симпатических нервных узлах, кроме свойственных для них мультиполярных нейронов, имеются группы мелких гранулосодержащпх интенсивно флуоресцирующих клеток (МИФ-клетки). Для них типичны короткие отростки и наличие в составе цитоплазмы гранулярных пузырьков. По флуоресценции и субмикроскопическому строению их пузырьки соответствуют пузырькам цитоплазмы клеток мозгового вещества надпочечников. Предполагается их участие в проведении нервного импульса от преганглионарных волокон на нейроны ганглия (роль интернейронов). МИФ-клетки рассматривают как внутриганглионарную тормозную систему. Возбуждаясь преганглионарными синапсами, они выделяют катехоламины, тормозящие передачу нервного импульса с преганглионарных волокон на симпатические нейроны узла.
Интрамуральные нервные сплетения. Значительное количество нейроцитов вегетативной нервной системы сосредоточено в нервных сплетениях полых органов: пищеварительного тракта, сердца, мочевого пузыря и др. Нервные узлы сплетений содержат эфферентные, рецепторные и ассоциативные нейроны. Морфологически в интрамуральных нервных узлах различают три типа нервных клеток (рис. 181).
Клетки первого типа Догеля (длинноаксонные невроциты) характеризуются длинным аксоном и многими ветвящимися дендритами. Клетки второго типа Догеля (равноотростчатые нейроциты) содержат несколько отростков, среди которых морфологически нельзя определить аксон. Экспериментально установлено, что он заканчивается синапсом на клетках первого типа. Клетки третьего типа образуют синаптические связи с дендритами нейронов соседних ганглиев. В стенке желудочно-кишечного тракта имеется три нервных сплетения: подсерозное, межмышечное и подслизистое, содержащие ганглии нервных клеток.
Наиболее массивное межмышечное нервное сплетение расположено в мышечной оболочке органа между продольным и циркулярным слоями. Нейрогистологически, электронно-микроскопически, гистохимически и нейрофизиологически выявлена специфичность этого сплетения, позволяющая сопоставить его по ряду признаков с центральной нервной системой. В частности, межмышечное нервное сплетение кишечника также покрыто соединительнотканной оболочкой, отграниченной от нервной ткани ба-зальной мембраной. Нервное сплетение имеет свою систему кровоснабжения в виде собственной капиллярной сети, локализованной за пределами капсулы. Капилляры и соединительная ткань в паренхиму ганглия не проникают.
Капсула сплетения содержит 2 — 3 слоя коллагеновых волокон, разграниченных друг от друга плоскими клетками. Внутри каждого слоя волокна ориентированы параллельно, не образуя пучков. В цитоплазме клеток капсулы различают свободные рибосомы, митохондрии и пиноцитозные пузырьки. Последние свидетельствуют об участии этих клеток в процессах транспорта веществ.
Экспериментами показано, что эндотелий капилляров и капсула сплетений участвуют в формировании барьера «кровь — нервное сплетение», препятствующего проникновению в последнее молекул маркеров.
Нейроглия ганглиев межмышечного нервного сплетения не дифференцирована (в отличие от экстрамуральных узлов) на сателлиты капсулы нейроцитов и леммоциты волокон. Глиоциты одновременно граничат с перикарпонами нейронов, покрывают группы осевых цилиндров и синаптические образования. Цитоплазма глиоцитов бедна органеллами. Они содержат небольшие цистерны гранулярной эндоплазматической сети, одиночные митохондрии и свободные полирибосомы.
Нейроциты ганглиев межмышечного нервного сплетения характеризуются обилием органелл. Крупные комплексы Гольджи и цистерны эндоплазматической сети в совокупности образуют в клетках плотную сеть мембран. Многочисленные рибосомы локализированы как свободно, так и на мембранах эндоплазматической сети. Гранулярная эндоплазматическая сеть распределена равномерно и не образует хроматофильных глыбок.
В ганглиях межмышечного нервного сплетения кишечника содержатся эфферентные и афферентные нейроны (рис. 182). Методом флуоресценции и электронной микроскопии на клетках первого типа Догеля выявляются возбуждающие холинергические и тормозные адренергические синапсы.
За небольшим исключением, интрамуральные нейроны кишечника неадренергические, но они снабжены адренергическими синапсами главным образом аксонов нервных клеток превертебральных ганглиев. Электронно-микроскопически эти синапсы характеризуются типичными для адренергических элементов гранулярными синаптическими пузырьками (30 — 60 нм).
Методом флуоресцентной микроскопии установлено, что адренергические синапсы имеют большинство нейронов сплетения. Отсутствие флуоресцирующих волокон при перерезке брыжеечных нервов свидетельствует, что адренергические синапсы в составе интрамуральных сплетений образованы окончаниями
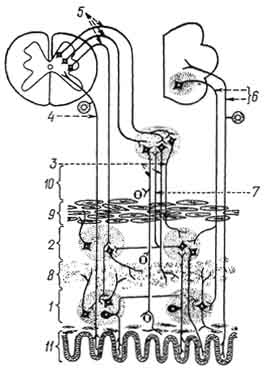
Рис. 182. Схема интрамуральных сплетений пищеварительного тракта:
1 — подслизистое сплетение; 2 — миэнтеральное сплетение; 3 — постганглионарные симпатические волокна; 4 — спинальные афферентные волокна; 5 — симпатические преганглионарные волокна; 6 — вагусные афферентные волокна; 7 — афферентные волокна местных рефлекторных путей; 8 — кольцевой мышечный слой; 9 — продольный мышечный слой; 10 — серозный слой; 11 — слизистая оболочка (по Ноздрачеву, 1978).
аксонов нейроцитов пре- или паравертебральных ганглиев.
В кишечнике, кроме холинергических нейроцитов, имеются тормозящие пуринергические нейроны, выделяющие в качестве медиатора пуриновые соединения. Перикарионы и их отростки содержат характерные для них электроноплотные пузырьки 100 нм в диаметре. Пуринергические нейроны вызывают нисходящее торможение пищеварительного тракта, что является звеном перистальтического рефлекса. В отличие от пуринергических адренергические нервы вызывают рефлекторное угнетение перистальтики торможением интрамуральных холинергических возбуждающих нейронов (Берсток и Коста, 1979).
Отзывов (0)
Добавить отзыв
Отделы вегетативной нервной системы. Строение вегетативной нервной системы.В вегетативной нервной системе различают центральные и периферические отделы. Центральные отделы симпатической нервной системы представлены ядрами боковых рогов тораколюмбального отдела спинного мозга. В парасимпатической нервной системе центральные отделы включают ядра среднего и продолговатого мозга, а также ядра боковых рогов сакрального отдела спинного мозга. Парасимпатические волокна краниобульбарного отдела выходят в составе III-й, VII-й, IX-й и Х-й пар черепных нервов. Вегетативные рефлекторные дуги начинаются чувствительным нейроном, тело которого лежит в спинномозговом узле (ганглии), как и в соматической рефлекторной дуг. Ассоциативные нейроны находятся в боковых рогах спинного мозга. Здесь нервные импульсы переключаются на промежуточные преганглионарные нейроны, отростки которых покидают центральные ядра и достигают вегетативных ганглиев, где передают импульсы на двигательный нейрон. В связи с этим различают нервные волокна преганглионарные и постганглионарные. Первые из них покидают центральную нервную систему в составе вентральных корешков спинномозговых нервов и черепных нервов. Как в симпатической, так и в парасимпатической системах преганглионарные нервные волокна принадлежат холинергическим нейронам. Аксоны нейронов, расположенных в вегетативных ганглиях, называются постганглионарными. Они не образуют прямых контактов с эффекторными клетками. Их терминальные отделы по своему ходу формируют расширения — варикозности, в составе которых находяся пузырьки медиатора. В области варикозности нет глиальной оболочки и нейромедиатор, выделяясь в окружающую среду, влияет на эффекторные клетки (например, на клетки желез, гладкие миоциты и др.). В периферических ганглиях симпатической нервной системы, как правило, находятся адренергические эфферентные нейроны (за исключением нейронов, имеющих синаптические связи с потовыми железами, где симпатические нейроны являются холинергическими). В парасимпатических ганглиях эфферентные нейроны всегда холинергические.
Ганглии представляют собой скопления мультиполярных нейронов (от нескольких клеток до десятков тысяч). Экстраорганные (симпатические) ганглии имеют хорошо выраженную соединительнотканную капсулу, как продолжение периневрия. Парасимпатические ганглии находятся, как правило, в интрамуральных нервных сплетениях. Ганглии интрамуральных сплетений, как и другие вегетативные узлы, содержат вегетативные нейроны местных рефлекторных дуг. Мультиполярные нейроны диаметром 20-35 мкм расположены диффузно, каждый нейрон окружен глиоцитами ганглия. Кроме того, описаны нейроэндокринные, хеморецепторные, биполярные, а у некоторых позвоночных и униполярные нейроны. В симпатических ганглиях имеются мелкие интенсивно флюоресцирующие клетки (МИФ-клетки) с короткими отростками и большим количеством гранулярных пузырьков в цитоплазме. Они выделяют катехоламины и оказывают тормозящее влияние на передачу импульсов с преганглионарных нервных волокон на эфферентный симпатический нейрон. Эти клетки называют интернейронами. Среди крупных мультиполярных нейронов вегетативных ганглиев различают: двигательные (клетки Догеля I-го типа), чувствительные (клетки Догеля II-го типа) и ассоциативные (клетки Догеля III-го типа). Двигательные нейроны имеют короткие дендриты с пластинчатыми расширениями («рецептивные площадки»). Аксон этих клеток очень длинный, уходит за пределы ганглия в составе постганглионарных тонких безмиелиновых нервных волокон и оканчивается на гладких миоцитах внутренних органов. Клетки I-го типа называют длинноаксонными нейронами. Нейроны II-го типа — равноотростчатые нервные клетки. От их тела отходят 2-4 отростка, среди которых различить аксон трудно. Не разветвляясь, отростки уходят далеко от тела нейрона. Их дендриты имеют чувствительные нервные окончания, а аксон оканчивается на телах двигательных нейронов в соседних ганглиях. Клетки II-го типа являются чувствительными нейронами местных вегетативных рефлекторных дуг. Клетки Догеля III-го типа по форме тела похожи на вегетативные нейроны П-го типа, но их дендриты не выходят за пределы ганглия, а нейрит направляется в другие ганглии. Многие исследователи считают эти клетки разновидностями чувствительных нейронов. Таким образом, в периферических вегетативных ганглиях имеются местные рефлекторные дуги, состоящие из чувствительных, двигательных и, возможно, ассоциативных вегетативных нейронов. Интрамуральные вегетативные ганглии в стенке пищеварительного тракта отличаются тем, что в их составе, кроме двигательных холинергических нейронов, имеются тормозные нейроны. Они представлены адренергическими и пуринергическими нервными клетками. В последних медиатором является пуриновый нуклеотид. В интрамуральных вегетативных ганглиях встречаются также пептидергические нейроны, выделяющие вазоинтестинальный пептид, соматостатин и ряд других пептидов, с помощью которых осуществляются нейроэндокринная регуляция и модуляция деятельности тканей и органов пищеварительной системы. Учебное видео анатомии вегетативной нервной системы (ВНС)При проблемах с просмотром скачайте видео со страницы Здесь — Также рекомендуем «Регенерация в нервной системе. Сенсорный комплекс органов.» Оглавление темы «Нервная система. Строение глаза.»: |
Симпатический (показан красным) и парасимпатический (показан синим) отделы автономной нервной системы
У этого термина существуют и другие значения, см. ВНС.
Вегетати́вная не́рвная систе́ма[1] (от лат. vegetatio — возбуждение, от лат. vegetativus — растительный), ВНС, автономная нервная система, ганглионарная нервная система (от лат. ganglion — нервный узел), висцеральная нервная система (от лат. viscera — внутренности), органная нервная система, чревная нервная система, systema nervosum autonomicum (PNA) — часть нервной системы организма, комплекс центральных и периферических клеточных структур, регулирующих функциональный уровень организма, необходимый для адекватной реакции всех его систем.
Вегетативная нервная система — отдел нервной системы, регулирующий деятельность внутренних органов, желез внутренней и внешней секреции, кровеносных и лимфатических сосудов[2]. Играет ведущую роль в поддержании постоянства внутренней среды организма и в приспособительных реакциях всех позвоночных.
Анатомически и функционально вегетативная нервная система подразделяется на симпатическую, парасимпатическую и метасимпатическую. Симпатические и парасимпатические центры находятся под контролем коры больших полушарий и гипоталамических центров[3].
В симпатическом и парасимпатическом отделах имеются центральная и периферическая части. Центральную часть образуют тела нейронов, лежащих в спинном и головном мозге. Эти скопления нервных клеток получили название вегетативных ядер. Отходящие от ядер волокна, вегетативные ганглии, лежащие за пределами центральной нервной системы, и нервные сплетения в стенках внутренних органов образуют периферическую часть вегетативной нервной системы.
Симпатические ядра расположены в спинном мозге. Отходящие от него нервные волокна заканчиваются за пределами спинного мозга в симпатических узлах, от которых берут начало нервные волокна. Эти волокна подходят ко всем органам.
Парасимпатические ядра лежат в среднем и продолговатом мозге и в крестцовой части спинного мозга. Нервные волокна от ядер продолговатого мозга входят в состав блуждающих нервов. От ядер крестцовой части нервные волокна идут к кишечнику, органам выделения.
Метасимпатическая нервная система представлена нервными сплетениями и мелкими ганглиями в стенках пищеварительного тракта, мочевого пузыря, сердца и некоторых других органов.
Деятельность вегетативной нервной системы не зависит от воли человека. Это означает, что в обычных условиях человек не может волевым усилием заставить сердце биться реже или мышцы желудка — не сокращаться. Однако достичь сознательного влияния на многие параметры, контролируемые ВНС, можно с помощью специальных методов тренировки — например, с использованием методов биологической обратной связи.
Симпатическая нервная система усиливает обмен веществ, повышает возбуждаемость большинства тканей, мобилизует силы организма на активную деятельность. Парасимпатическая система способствует восстановлению израсходованных запасов энергии, регулирует работу организма во время сна.
Под контролем автономной системы находятся органы кровообращения, дыхания, пищеварения, выделения, размножения, а также обмен веществ и рост. Фактически эфферентный отдел ВНС осуществляет нервную регуляцию функций всех органов и тканей, кроме скелетных мышц, которыми управляет соматическая нервная система.
В отличие от соматической нервной системы, двигательный эффекторный нейрон в автономной нервной системе находится на периферии, и спинной мозг лишь косвенно управляет его импульсами.
Термины автономная система, висцеральная система, симпатический отдел нервной системы неоднозначны. В настоящее время симпатическими называют только часть висцеральных эфферентных волокон. Однако различные авторы используют термин «симпатический» по-разному:
- в узком понимании, как описано в предложении выше;
- в качестве синонима термина «автономный»;
- как название всей висцеральной («вегетативной»)[4] нервной системы — как афферентной, так и эфферентной.
Терминологическая путаница возникает также, когда автономной называют всю висцеральную систему (и афферентную, и эфферентную).
Классификация отделов висцеральной нервной системы позвоночных, приведённая в руководстве[5] А. Ромера и Т. Парсонса, выглядит следующим образом:
Висцеральная нервная система:
- афферентная;
- эфферентная:
- особая жаберная;
- автономная:
- симпатическая;
- парасимпатическая.
Морфология[править | править код]
Выделение автономной (вегетативной) нервной системы обусловлено некоторыми особенностями её строения. К этим особенностям относятся следующие:
- очаговость локализации вегетативных ядер в ЦНС;
- скопление тел эффекторных нейронов в виде узлов (ганглиев) в составе вегетативных сплетений;
- двухнейронность нервного пути от вегетативного ядра в ЦНС к иннервируемому органу.
Волокна автономной нервной системы выходят не сегментарно, как в соматической нервной системе, а из трёх отстоящих друг от друга ограниченных участков мозга: черепного, грудинопоясничного и крестцового.
Автономную нервную систему разделяют на симпатическую, парасимпатическую и метасимпатическую части. В симпатической части отростки спинномозговых нейронов короче, ганглионарные длиннее. В парасимпатической системе, наоборот, отростки спинномозговых клеток длиннее, ганглионарных короче. Симпатические волокна иннервируют все без исключения органы, в то время как область иннервации парасимпатических волокон более ограничена.
Центральный и периферический отделы[править | править код]
Автономная (вегетативная) нервная система подразделяется по топографическому признаку делятся на центральный и периферический отделы.
Центральный отдел[править | править код]
- парасимпатические ядра 3, 7, 9 и 10 пар черепных нервов, лежащие в мозговом стволе (краниобульбарный отдел).;ядра, лежащие в среднем мозге(мезоенцефалический отдел).; ядра, залегающие в сером веществе трёх крестцовых сегментов (сакральный отдел)[6]
- симпатические ядра, расположенные в боковых рогах тораколюмбального отдела спинного мозга;
Периферический отдел[править | править код]
- вегетативные (автономные) нервы, ветви и нервные волокна, выходящие из головного и спинного мозга;
- вегетативные (автономные, висцеральные) сплетения;
- узлы (ганглии) вегетативных (автономных, висцеральных) сплетений;
- симпатический ствол (правый и левый) с его узлами (ганглиями), межузловыми и соединительными ветвями и симпатическими нервами;
- концевые узлы (ганглии) парасимпатической части вегетативной нервной системы.
Симпатический, парасимпатический и метасимпатический отделы[править | править код]
На основании топографии вегетативных ядер и узлов, различий в длине аксонов первого и второго нейронов эфферентного пути, а также особенностей функции вегетативная нервная система подразделяется на симпатическую, парасимпатическую и метасимпатическую.
Расположение ганглиев и строение проводящих путей[править | править код]
Нейроны ядер центрального отдела вегетативной нервной системы — первые эфферентные нейроны на пути от ЦНС (спинной и головной мозг) к иннервируемому органу. Нервные волокна, образованные отростками этих нейронов, носят название предузловых (преганглионарных) волокон, так как они идут до узлов периферической части вегетативной нервной системы и заканчиваются синапсами на клетках этих узлов. Преганглионарные волокна имеют миелиновую оболочку, благодаря чему отличаются беловатым цветом. Они выходят из мозга в составе корешков соответствующих черепных нервов и передних корешков спинномозговых нервов.
Вегетативные узлы (ганглии): входят в состав симпатических стволов (есть у большинства позвоночных, кроме круглоротых и хрящевых рыб), крупных вегетативных сплетений брюшной полости и таза, располагаются в области головы и в толще или возле органов пищеварительной и дыхательной систем, а также мочеполового аппарата, которые иннервируются вегетативной нервной системой. Узлы периферической части вегетативной нервной системы содержат тела вторых (эффекторных) нейронов, лежащих на пути к иннервируемым органам. Отростки этих вторых нейронов эфферентного пути, несущих нервный импульс из вегетативных узлов к рабочим органам (гладкая мускулатура, железы, ткани), являются послеузелковыми (постганглионарными) нервными волокнами. Из-за отсутствия миелиновой оболочки они имеют серый цвет. Постганглионарные волокна автономной нервной системы в большинстве своем тонкие (чаще всего их диаметр не превышает 7 мкм) и не имеют миелиновой оболочки. Поэтому возбуждение по ним распространяется медленно, а нервы автономной нервной системы характеризуются бо́льшим рефрактерным периодом и большей хронаксией.
Рефлекторная дуга[править | править код]
Строение рефлекторных дуг вегетативного отдела отличается от строения рефлекторных дуг соматической части нервной системы. В рефлекторной дуге вегетативной части нервной системы эфферентное звено состоит не из одного нейрона, а из двух, один из которых находится вне ЦНС. В целом простая вегетативная рефлекторная дуга представлена тремя нейронами.
Первое звено рефлекторной дуги — это чувствительный нейрон, тело которого располагается в спинномозговых узлах и в чувствительных узлах черепных нервов. Периферический отросток такого нейрона, имеющий чувствительное окончание — рецептор, берёт начало в органах и тканях. Центральный отросток в составе задних корешков спинномозговых нервов или чувствительных корешков черепных нервов направляется к соответствующим ядрам в спинной или головной мозг.
Второе звено рефлекторной дуги является эфферентным, поскольку несёт импульсы из спинного или головного мозга к рабочему органу. Этот эфферентный путь вегетативной рефлекторной дуги представлен двумя нейронами. Первый из этих нейронов, второй по счёту в простой вегетативной рефлекторной дуге, располагается в вегетативных ядрах ЦНС. Его можно называть вставочным, так как он находится между чувствительным (афферентным) звеном рефлекторной дуги и вторым (эфферентным) нейроном эфферентного пути.
Эффекторный нейрон представляет собой третий нейрон вегетативной рефлекторной дуги. Тела эффекторных (третьих) нейронов лежат в периферических узлах вегетативной нервной системы (симпатический ствол, вегетативные узлы черепных нервов, узлы внеорганных и внутриорганных вегетативных сплетений). Отростки этих нейронов направляются к органам и тканям в составе органных вегетативных или смешанных нервов. Заканчиваются постганглионарные нервные волокна на гладких мышцах, железа́х и в других тканях соответствующими концевыми нервными аппаратами.
Физиология[править | править код]
Общее значение вегетативной регуляции[править | править код]
Вегетативная нервная система приспосабливает работу внутренних органов к изменениям окружающей среды. ВНС обеспечивает гомеостаз (постоянство внутренней среды организма). ВНС также участвует во многих поведенческих актах, осуществляемых под управлением головного мозга, влияя не только на физическую, но и на психическую деятельность человека.
Роль симпатического и парасимпатического отделов[править | править код]
Симпатическая нервная система активируется при стрессовых реакциях. Для неё характерно генерализованное влияние, при этом симпатические волокна иннервируют подавляющее большинство органов.
Известно, что парасимпатическая стимуляция одних органов оказывает тормозное действие, а других — возбуждающее действие. В большинстве случаев действие парасимпатической и симпатической систем противоположно.
Влияние симпатического и парасимпатического отделов на отдельные органы[править | править код]
Влияние симпатического отдела:
- На сердце — повышает частоту и силу сокращений сердца.
- На артерии —[7]сужает артерии большинства органов, расширяет артерии скелетных мышц.
- На кишечник — угнетает перистальтику кишечника и выработку пищеварительных ферментов.
- На слюнные железы — угнетает слюноотделение.
- На мочевой пузырь — расслабляет мочевой пузырь.
- На бронхи и дыхание — расширяет бронхи и бронхиолы, усиливает вентиляцию лёгких.
- На зрачок — расширяет зрачки.
Влияние парасимпатического отдела:
- На сердце — уменьшает частоту и силу сокращений сердца.
- На артерии — не влияет в большинстве органов, вызывает расширение артерий половых органов и мозга, сужение коронарных артерий и артерий лёгких.
- На кишечник — усиливает перистальтику кишечника и стимулирует выработку пищеварительных ферментов.
- На слюнные железы — стимулирует слюноотделение.
- На мочевой пузырь — сокращает мочевой пузырь.
- На бронхи и дыхание — сужает бронхи и бронхиолы, уменьшает вентиляцию лёгких.
- На зрачок — сужает зрачки.
Нейромедиаторы и клеточные рецепторы[править | править код]
Симпатический и парасимпатический отделы оказывают различное, в ряде случаев противонаправленное влияние на различные органы и ткани, а также перекрёстно влияют друг на друга. Различное воздействие этих отделов на одни и те же клетки связано со спецификой выделяемых ими нейромедиаторов и со спецификой рецепторов, имеющихся на пресинаптических и постсинаптических мембранах нейронов автономной системы и их клеток-мишеней.
Преганглионарные нейроны обоих отделов автономной системы в качестве основного нейромедиатора выделяют ацетилхолин, который действует на никотиновые рецепторы ацетилхолина на постсинаптической мембране постганглионарных (эффекторных) нейронов. Постганглионарные нейроны симпатического отдела, как правило, выделяют в качестве медиатора норадреналин, который действует на адренорецепторы клеток-мишеней. На клетках-мишенях симпатических нейронов бета-1 и альфа-1 адренорецепторы в основном сосредоточены на постсинаптических мембранах (это означает, что in vivo на них действует в основном норадреналин), а аль-2 и бета-2 рецепторы — на внесинаптических участках мембраны (на них в основном действует адреналин крови). Лишь некоторые постганглионарные нейроны симпатического отдела (например, действующие на потовые железы) выделяют ацетилхолин.
Постганглионарные нейроны парасимпатического отдела выделяют ацетилхолин, который действует на мускариновые рецепторы клеток-мишеней.
На пресинаптической мембране постганглионарных нейронов симпатического отдела преобладают два типа адренорецепторов: альфа-2 и бета-2 адренорецепторы. Кроме того, на мебране этих нейронов расположены рецепторы к пуриновым и пиримидиновым нуклеотидоам (P2X-рецепторы АТФ и др.), никотиновые и мускариновые холинорецепторы, рецепторы нейропептидов и простагландинов, опиоидные рецепторы[8].
При действии на альфа-2 адренорецепторы норадреналина или адреналина крови падает внутриклеточная концентрация ионов Ca2+, и выделение норадреналина в синапсах блокируется. Возникает петля отрицательной обратной связи. Альфа-2 рецепторы более чувствительны к норадреналину, чем к адреналину.
При действии норадреналина и адреналина на бета-2 адренорецепторы выделение норадреналина обычно усиливается. Этот эффект наблюдается при обычном взаимодействии с Gs-белком, при котором растёт внутриклеточная концентрация цАМФ. Бета-два рецепторы более чувствительны к адреналину. Поскольку под действием норадреналина симпатических нервов из мозгового слоя надпочечников выделяется адреналин, возникает петля положительной обратной связи.
Однако в некоторых случаях активация бета-2 рецепторов может блокировать выделение норадреналина. Показано, что это может быть следствием взаимодействия бета-2 рецепторов с Gi/o белками и связывания (секвестирования) ими Gs-белков, которое, в свою очередь, предотвращает взаимодействие Gs-белков с другими рецепторами [1].
При действии ацетилхолина на мускариновые рецепторы симпатических нейронов выделение норадреналина в их синапсах блокируется, а при действии на никотиновые рецепторы — стимулируется. Поскольку на пресинаптических мембранах симпатических нейронов преобладают мускариновые рецепторы, обычно активация парасимпатических нервов снижает уровень выделения норадреналина из симпатических нервов.
На пресинаптических мембранах постганглионарных нейронов парасимпатического отдела преобладают альфа-2 адренорецепторы. При действии на них норадреналина выделение ацетилхолина блокируется. Таким образом, симпатические и парасимпатические нервы взаимно ингибируют друг друга.
Развитие в эмбриогенезе[править | править код]
- Развитие периферической (соматической) и вегетативной нервной системы. Периферическая (соматическая) и вегетативная нервная система развивается из наружного зародышевого листка — эктодермы. Черепные и спинномозговые нервы у плода закладываются очень рано (5-6 нед). Миелинизация нервных волокон происходит позже (у преддверного нерва — 4 мес; у большинства нервов — на 6-7-м месяце).
Спинномозговые и периферические вегетативные узлы закладываются одновременно с развитием спинного мозга. Исходным материалом для них служат клеточные элементы ганглиозной пластинки, её нейробласты и глиобласты, из которых образуются клеточные элементы спинномозговых узлов. Часть их смещается на периферию в места локализации вегетативных нервных узлов
Сравнительная анатомия и эволюция вегетативной нервной системы[править | править код]
У насекомых имеется так называемая симпатическая, или стомодеальная нервная система[9]. В её состав входит фронтальный ганглий, который находится спереди от головного мозга и соединён парными коннективами с тритоцеребрумом. От него отходит непарный фронтальный нерв, тянущийся вдоль спинной стороны глотки и пищевода. Этот нерв соединяется с несколькими нервными ганглиями; отходящие от них нервы иннервируют переднюю кишку, слюнные железы и аорту.
См. также[править | править код]
- Симпатическая нервная система
- Парасимпатическая нервная система
- Метасимпатическая нервная система
- Соматическая нервная система
- Центральная нервная система (ЦНС)
- Индекс Кердо
- Индекс Баевского
Примечания[править | править код]
- ↑ Лапин C. К., Михайловский В. С., Плечкова Е. К., Шефер Д. Г. Вегетативная нервная система // Большая медицинская энциклопедия, 3-е изд. — М.: Советская энциклопедия. — Т. 4.
- ↑ Краткая медицинская энциклопедия.
- ↑ Словарь биолога. (недоступная ссылка)
- ↑ Например, в книге «Физиология человека / Под ред. В. М. Покровского, Г. Ф. Коротько. — М.: Медицина, 1997. — Т. 1 — 448 с.; Т. 2 — 368 с.».
- ↑ Ромер А., Парсонс Т. Анатомия позвоночных. — Т. 2. — С. 260.
- ↑ I. Espinosa-Medina, O. Saha, F. Boismoreau, Z. Chettouh, F. Rossi. The sacral autonomic outflow is sympathetic (англ.) // Science. — 2016-11-18. — Vol. 354, iss. 6314. — P. 893–897. — ISSN 1095-9203 0036-8075, 1095-9203. — doi:10.1126/science.aah5454.
- ↑ Брин В. Б. и др. Основы физиологии человека в 2-х т. Учебник для высших учебных заведений. Ред. Б. И. Ткаченко. СПб., 1994. Т.1 — 567 с. Т.2 — 413 с.
- ↑ Stefan Boehm, Sigismund Huck. Receptors controlling transmitter release from sympathetic neurons in vitro // Progress in Neurobiology. — Volume 51, Issue 3, February 1997, Pages 225—242. (недоступная ссылка)
- ↑ Г. Росс, Ч. Росс, Д. Росс. Энтомологи?