Какие конечные продукты подготовительного этапа энергетического обмена


СТАДИИ ЭНЕРГЕТИЧЕСКОГО ОБМЕНА
Энергетический обмен (катаболизм, диссимиляция) — это процессы расщепления ве ществ с высвобождением энергии. Высвобожденная энергия преобразуется в энергию АТФ. Наиболее важными процессами энергетического обмена являются дыхание и брожение.

Энергетический обмен – это совокупность химических реакций постепенного распада органических соединений, сопровождающихся высвобождением энергии, часть которой расходуется на синтез АТФ. Синтезированная АТФ становится универсальным источником энергии для жизнедеятельности организмов. Она образуется в результате реакции фосфорилирования – присоединения остатков фосфорной кислоты к молекуле АДФ. На эту реакцию расходуется энергия, которая затем накапливается в макроэргических связях молекулы АТФ, при распаде молекулы АТФ или при ее гидролизе до АДФ клетка получает около 40 кДж энергии.
АТФ – постоянный источник энергии для клетки, она мобильно может доставлять химическую энергию в любую часть клетки. Когда клетке необходима энергия – достаточно гидролизовать молекулу АТФ. Энергия выделяется в результате реакции диссимиляции (расщепления органических веществ), в зависимости от специфики организма и условий его обитания энергетический обмен проходит в два или три этапа. Большинство живых организмов относятся к аэробам, использующим для обмена веществ кислород, который поступает из окружающей среды. Для аэробов энергетический обмен проходит в три этапа:
— подготовительный;
— бескислородный;
— кислородный.
В организмах, которые обитают в бескислородной среде и не нуждаются в кислороде для энергетического обмена – анаэробах и аэробах, при недостатке кислорода проходят энергетический обмен в два этапа:
— подготовительный;
— бескислородный.
Количество энергии, которое выделяется при двухэтапном варианте намного меньше, чем в трехэтапном.
ЭТАПЫ ЭНЕРГЕТИЧЕСКОГО ОБМЕНА
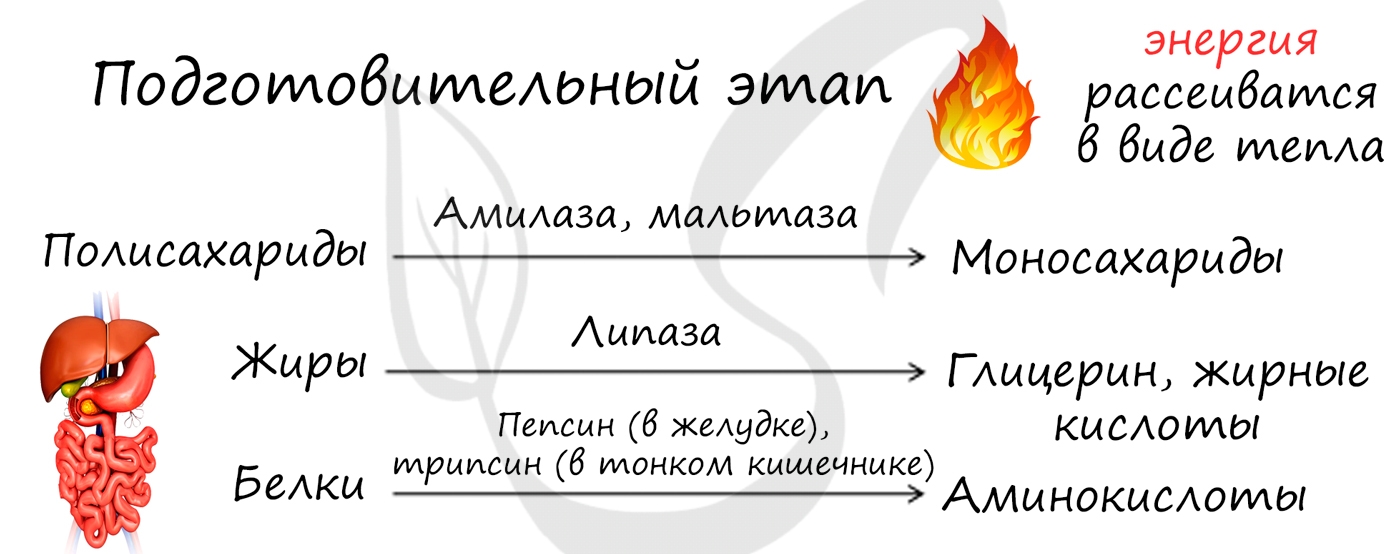
Подготовительный этап – во время него крупные пищевые полимерные молекулы распадаются на более мелкие фрагменты. В желудочно-кишечном тракте многоклеточных организмов он осуществляется пищеварительными ферментами, у одноклеточных – ферментами лизосом. Полисахариды распадаются на ди- и моносахариды, белки – до аминокислот, жиры – до глицерина и жирных кислот. В ходе этих превращений энергии выделяется мало, она рассеивается в виде тепла, и АТФ не образуется. Образующиеся в ходе подготовительного этапа соединения-мономеры могут участвовать в реакциях пластического обмена (в дальнейшем из них синтезируются вещества, необходимые для клетки) или подвергаться дальнейшему расщеплению с целью получения энергии.
Большинство клеток в первую очередь используют углеводы, жиры остаются в первом резерве и используются по окончания запаса углеводов. Хотя есть и исключения: в клетках скелетных мышц при наличии жирных кислот и глюкозы предпочтение отдается жирным кислотам. Белки расходуются в последнюю очередь, когда запас углеводов и жиров будет исчерпан – при длительном голодании.
Бескислородный этап (гликолиз) – происходит в цитоплазме клеток. Главным источником энергии в клетке является глюкоза. Ее бескислородное расщепление называют анаэробным гликолизом. Он состоит из ряда последовательных реакций по превращению глюкозы в лактат. Его присутствие в мышцах хорошо известно уставшим спортсменам. Этот этап заключается в ферментативном расщеплении органических веществ, полученных в ходе первого этапа. Так как глюкоза является наиболее доступным субстратом для клетки как продукт расщепления полисахаридов, то второй этап можно рассмотреть на примере ее бескислородного расщепления – гликолиза (Рис. 1).
Рис. 1. Бескислородный этап
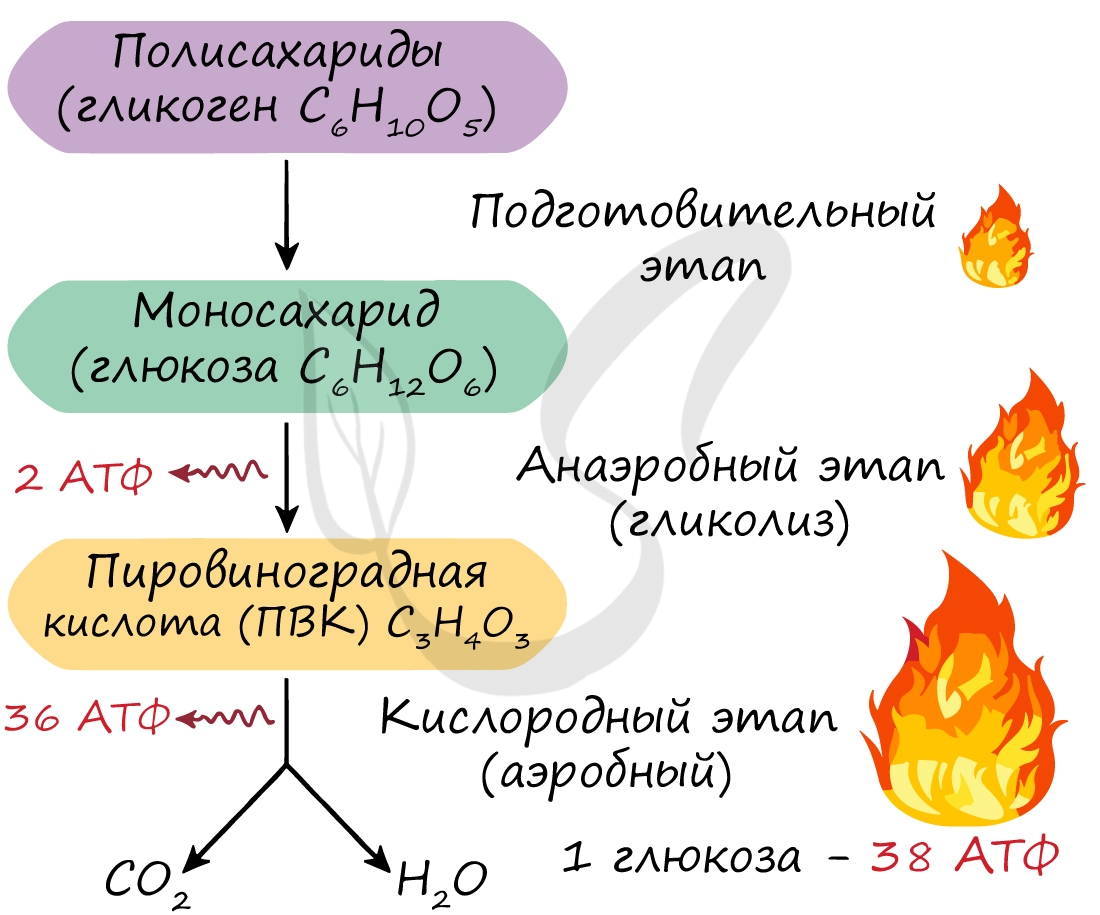
Гликолиз – многоступенчатый процесс бескислородного расщепления молекулы глюкозы, содержащей шесть атомов углерода, до двух молекул пировиноградной кислоты (пируват). Реакция гликолиза катализируется многими ферментами и протекает в цитоплазме клетки. В ходе гликолиза при расщеплении одного моля глюкозы выделяется около 200 кДж энергии, 60 % ее рассеивается в виде тепла, 40 % – для синтезирования двух молекул АТФ из двух молекул АДФ. При наличии кислорода в среде пировиноградная кислота из цитоплазмы переходит в митохондрии и участвует в третьем этапе энергетического обмена. Если кислорода в клетке нет, то пировиноградная кислота преобразуется в животных клетках или превращается в молочную кислоту.
В микроорганизмах, которые существуют без доступа кислорода – получают энергию в процессе брожения, начальный этап аналогичен гликолизу: распад глюкозы до двух молекул пировиноградной кислоты, и далее она зависит от ферментов, которые находятся в клетке – пировиноградная кислота может преобразовываться в спирт, уксусную кислоту, пропионовую и молочную кислоту. В отличие от того, что происходит в животных тканях, у микроорганизмов этот процесс носит название молочнокислого брожения. Все продукты брожения широко используются в практической деятельности человека: это вино, квас, пиво, спирт, кисломолочные продукты. При брожении, так же как и при гликолизе, выделяется всего две молекулы АТФ.
Кислородный этап стал возможен после накопления в атмосфере достаточного количества молекулярного кислорода, он происходит в митохондриях клеток. Он очень сложен по сравнению с гликолизом, это процесс многостадийный и идет при участии большого количества ферментов. В результате третьего этапа энергетического обмена из двух молекул пировиноградной кислоты формируется углекислый газ, вода и 36 молекул АТФ (Рис. 2).
Рис. 2. Митохондрия
Две молекулы АТФ запасаются в ходе бескислородного расщепления молекулами глюкозы, поэтому суммарный энергетический обмен в клетке в случае распада глюкозы можно представить как:
С6Н12О6 + 6О2 + 38АДФ + 38Н3РО4 = 6СО2 + 44Н2О + 38АТФ
В результате окисления одной молекулы глюкозы шестью молекулами кислорода образуется шесть молекул углекислого газа и выделяется тридцать восемь молекул АТФ.
Мы видим, что в трехэтапном варианте энергетического обмена выделяется гораздо больше энергии, чем в двухэтапном варианте – 38 молекул АТФ против 2.


БРОЖЕНИЕ

В отсутствие кислорода или при его недостатке про исходит брожение. Брожение является эволюционно бо лее ранним способом получения энергии, чем дыхание, однако оно энергетически менее выгодно, поскольку ко нечными продуктами брожения являются органические вещества, богатые энергией. Существует несколько видов брожения, названия которых определяются конечными продуктами: молочнокислое, спиртовое, уксуснокислое и др. Так, в скелетных мышцах в отсутствие кислорода протекает молочнокислое брожение, в ходе которого пировиноградная кислота восстанавли вается до молочной кислоты. При этом восстановленные ранее коферменты НАДН расходу ются на восстановление пирувата:

Энергетическая эффективность молочнокислого брожения составляет две молекулыАТФ, образованные в процессе окисления глюкозы до пирувата.
Для многих микроорганизмов брожение является единственным способом ассимиляции энергии. Большинство таких организмов живет в анаэробных условиях и погибает в присутствии кислорода, но есть и такие, которые нормально существуют и в присутствии кислорода, и без него. Например, дрожжевые грибы при спиртовом брожении окисляют пировиноградную кислоту до этилового спирта и оксида углерода (IV):


ВИДЕО ДОМА
Вопросы части с

Универсальным источником энергии во всех клетках служит АТФ (аденозинтрифосфат, или аденозинтрифосфорная кислота).
Все энергетические затраты любой клетки обеспечиваются за счёт универсального энергетического вещества — АТФ.
АТФ синтезируется в результате реакции фосфорилирования, то есть присоединения одного остатка фосфорной кислоты к молекуле АДФ (аденозиндифосфата):
АДФ + H3PO4+ 40 кДж = АТФ + H2O.
Энергия запасается в форме энергии химических связей АТФ. Химические связи АТФ, при разрыве которых выделяется много энергии, называются макроэргическими.
При распаде АТФ до АДФ клетка за счёт разрыва макроэргической связи получит приблизительно (40) кДж энергии.
Энергия для синтеза АТФ из АДФ выделяется в процессе диссимиляции.
Энергетический обмен (диссимиляция, катаболизм) — это совокупность химических реакций постепенного распада органических соединений, сопровождающихся высвобождением энергии, часть которой расходуется на синтез АТФ.
В зависимости от среды обитания организма, диссимиляция может проходить в два или в три этапа.
Процессы расщепления органических соединений у аэробных организмов происходят в три этапа: подготовительный, бескислородный и кислородный.
В результате этого органические вещества распадаются до простейших неорганических соединений.
У анаэробных организмов, обитающих в бескислородной среде и не нуждающихся в кислороде (а также у аэробных организмов при недостатке кислорода), диссимиляция происходит в два этапа: подготовительный и бескислородный.
В двухэтапном энергетическом обмене энергии запасается гораздо меньше, чем в трёхэтапном.
Первый этап — подготовительный
Подготовительный этап заключается в распаде крупных органических молекул до более простых: полисахаридов — до моносахаридов, липидов — до глицерина и жирных кислот, белков — до аминокислот.
Этот процесс называется пищеварением. У многоклеточных организмов он осуществляется в желудочно-кишечном тракте с помощью пищеварительных ферментов. У одноклеточных организмов — происходит под действием ферментов лизосом.
В ходе биохимических реакций, происходящих на этом этапе, энергии выделяется мало, она рассеивается в виде тепла, и АТФ не образуется.
Второй этап — бескислородный (гликолиз)
Второй (бескислородный) этап заключается в ферментативном расщеплении органических веществ, которые были получены в ходе подготовительного этапа. Кислород в реакциях этого этапа не участвует.
Биологический смысл второго этапа заключается в начале постепенного расщепления и окисления глюкозы с накоплением энергии в виде (2) молекул АТФ.
Процесс бескислородного расщепления глюкозы называется гликолиз.
Гликолиз происходит в цитоплазме клеток.
Он состоит из нескольких последовательных реакций превращения молекулы глюкозы C6H12O6 в две молекулы пировиноградной кислоты — ПВК C3H4O3 и две молекулы АТФ (в виде которой запасается примерно (40) % энергии, выделившейся при гликолизе). Остальная энергия (около (60) %) рассеивается в виде тепла.
C6H12O6+2H3PO4+2АДФ=2C3H4O3+2АТФ +2H2O.
Получившаяся пировиноградная кислота при недостатке кислорода в клетках животных, а также клетках многих грибов и микроорганизмов, превращается в молочную кислоту C3H6O3.
HOOC−CO−CH3пировиноградная кислота→НАД⋅H+H+лактатдегидрогеназаHOOC−CHOH−CH3молочная кислота.
В мышцах человека при больших нагрузках и нехватке кислорода образуется молочная кислота и появляется боль. У нетренированных людей это происходит быстрее, чем у людей тренированных.
При недостатке кислорода в клетках растений, а также в клетках некоторых грибов (например, дрожжей), вместо гликолиза происходит спиртовое брожение: пировиноградная кислота распадается на этиловый спирт C2H5OH и углекислый газ CO2:
C6H12O6+2H3PO4+2АДФ=2C2H5OH+2CO2+2АТФ+2H2O.
Третий этап — кислородный
В результате гликолиза глюкоза распадается не до конечных продуктов (CO2 и H2O), а до богатых энергией соединений (молочная кислота, этиловый спирт) которые, окисляясь дальше, могут дать её в больших количествах. Поэтому у аэробных организмов после гликолиза (или спиртового брожения) следует третий, завершающий этап энергетического обмена — полное кислородное расщепление, или клеточное дыхание.
Этот этап происходит на кристах митохондрий.
Третий этап, так же как и гликолиз, является многостадийным и состоит из двух последовательных процессов — цикла Кребса и окислительного фосфорилирования.
Третий (кислородный) этап заключается в том, что при кислородном дыхании ПВК окисляется до окончательных продуктов — углекислого газа и воды, а энергия, выделяющаяся при окислении, запасается в виде (36) молекул АТФ ((2) молекулы в цикле Кребса и (34) молекулы в ходе окислительного фосфорилирования).
Этот этап можно представить себе в следующем виде:
2C3H4O3+6O2+36H3PO4+36АДФ=6CO2+42H2O+36АТФ.
Вспомним, что ещё две молекулы АТФ запасаются в ходе бескислородного расщепления каждой молекулы глюкозы (на втором, бескислородном, этапе). Таким образом, в результате полного расщепления одной молекулы глюкозы образуется (38) молекул АТФ.
Суммарная реакция энергетического обмена:
C6H12O6+6O2=6CO2+6H2O+38АТФ.
Для получения энергии в клетках, кроме глюкозы, могут быть использованы и другие вещества: липиды, белки. Однако ведущая роль в энергетическом обмене у большинства организмов принадлежит сахарам.
Источники:
Каменский А. А., Криксунов Е. А., Пасечник В. В. Биология. 9 класс // ДРОФА.
Каменский А. А., Криксунов Е. А., Пасечник В. В. Биология. Общая биология (базовый уровень) 10–11 класс // ДРОФА.
Лернер Г. И. Биология: Полный справочник для подготовки к ЕГЭ: АСТ, Астрель.
Обмен веществ
Обмен веществ (метаболизм) складывается из процессов расщепления и синтеза — диссимиляции и ассимиляции, постоянно
протекающих в организме. Чтобы жизнь продолжалась, количество поступающей энергии должно превышать (или как минимум равняться)
количеству расходуемой энергии, поэтому диссимиляция и ассимиляция поддерживают определенный баланс друг с другом.

Энергетический обмен
Энергетический обмен (диссимиляция — от лат. dissimilis ‒ несходный) — обратная ассимиляции сторона обмена веществ, совокупность реакций, которые приводят к высвобождению энергии химических связей. Это реакции расщепления жиров,
белков, углеводов, нуклеиновых кислот до простых веществ.
Возможно три этапа диссимиляции: подготовительный, анаэробный и аэробный. Среда обитания определяет количество
этапов диссимиляции. Их может быть три, если организм обитает в кислородной среде, и два, если речь идет об
организме, обитающем в бескислородной среде (к примеру, в кишечнике).
Обсудим этапы энергетического обмена более подробно:
- Подготовительный этап
- Бескислородный этап (анаэробный) — гликолиз
- Кислородный этап (аэробный)
Осуществляется в ферментами, в результате действия которых, сложные вещества превращаются в более простые: полимеры распадаются на мономеры. Это сопровождается разрывом химических связей и выделением энергии, большая часть
которой рассеивается в виде тепла.
Под действием ферментов белки расщепляются на аминокислоты, жиры — на глицерин и жирные кислоты, сложные углеводы — до простых сахаров.
Этот этап является последним для организмов-анаэробов, обитающих в условиях, где кислород отсутствует. На этапе гликолиза
происходит расщепление молекулы глюкозы: образуется 2 молекулы АТФ и 2 молекулы пировиноградной кислоты (ПВК).
Происходит данный этап в цитоплазме клеток.
Этот этап доступен только для аэробов — организмов, живущих в кислородной среде. Из каждой молекулы ПВК, образовавшейся на
этапе гликолиза, синтезируется 18 молекул АТФ — в сумме с двух ПВК выход составляет 36 молекул АТФ.
Таким образом, суммарно с одной молекулы глюкозы можно получить 38 АТФ (гликолиз + кислородный этап).
Кислородный этап протекает на кристах митохондрий (складках, выпячиваниях внутренней мембраны), где наибольшая концентрация окислительных ферментов. Главную роль в этом процессе играет так называемый цикл Кребса, который подробно изучает биохимия.
АТФ — аденозинтрифосфорная кислота
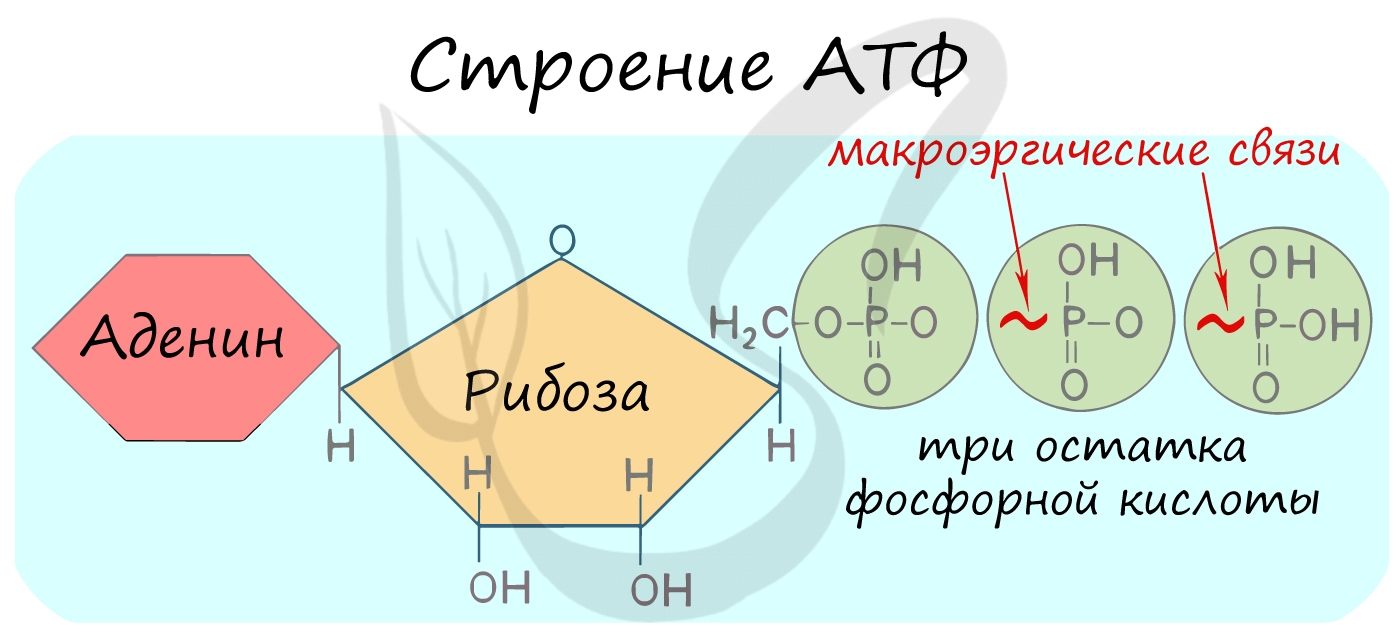
Трудно переоценить роль в клетке АТФ — универсального источника энергии. Молекула АТФ состоит из азотистого основания —
аденина, углевода — рибозы и трех остатков фосфорной кислоты.
Между остатками фосфорной кислоты находятся макроэргические связи — ковалентные связи, которые гидролизуются с выделением
большого количества энергии. Их принято обозначать типографическим знаком тильда «∽».
АТФ гидролизуется до АДФ (аденозиндифосфорная кислота), а затем и до АМФ (аденозинмонофосфорная кислота).
Гидролиз АТФ сопровождается выделением энергии (E) на каждом этапе и может быть представлен такой схемой:
- АТФ + H2O = АДФ + H3PO4 + E
- АДФ + H2O = АМФ + H3PO4 + E
- АМФ + H2O = аденин + рибоза + H3PO4 + E
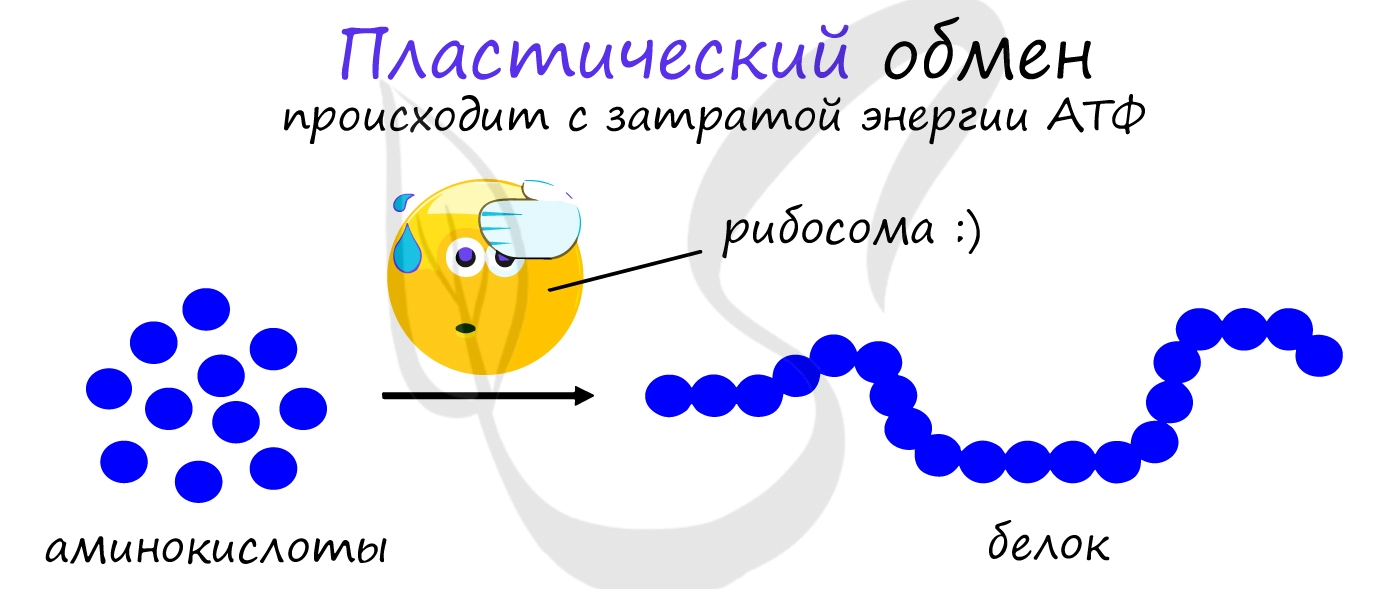
Пластический обмен
АТФ является универсальным источником энергии в клетке: энергия макроэргических связей АТФ используется для реакций
пластического обмена (ассимиляции), протекающих с затратой энергии: синтеза белка на рибосоме (трансляции),
удвоению ДНК (репликации) и т.д.
В результате пластического обмена в нашем организме происходит синтез белков, жиров и углеводов.
© Беллевич Юрий Сергеевич 2018-2020
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение
(в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов
без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования,
обратитесь, пожалуйста, к Беллевичу Юрию.
Этапы энергетического обмена
Значение АТФ в обмене веществ.
Хотя автотрофы и гетеротрофы получают энергию из внешней среды в различных формах, оба эти типа клеток накапливают и используют ее в основном
в одной форме — в виде аденозинтрифосфорной кислоты (АТФ).
Аденозинтрифосфорная кислота (АТФ) — мононуклеотид, содержащий азотистое основание аденин, 5-углеродный сахар рибозу и три остатка фосфорной
кислоты.
Ковалентные связи, соединяющие две фосфатные группы и содержащие большое количество энергии, называются
высокоэнергетическими(макроэргическими).
Молекула АТФ обладает способностью отдавать и присоединять остаток фосфорной кислоты (дефосфорилирование — фосфорилирование)
Таким образом, система АТФ/АДФ служит связующим звеном, объединяющим процессы катаболизма и анаболизма.
В реакциях катаболизма освобождающаяся химическая энергия используется
для фосфорилирования низкоэнергетического АДФ с превращением его в высокоэнергетический АТФ.
АТФ служит универсальным переносчиком и основным аккумулятором энергии в клетке.
При гидролизе под действием ферментов концевая фосфатная группа отщепляется от молекулы АТФ, в результате чего образуется молекула
аденозиндифосфата и выделяется 30,6 кДж энергии:
АТФ + Н20 → АДФ +Н3Р04 + 30,6 кДж.
Энергия, высвобождающаяся при гидролизе АТФ (обычно до стадии АДФ), расходуется клеткой для совершения всех видов работ: биологического синтеза
сложных органических соединений, транспорта веществ, движения (включая мышечное сокращение) и др.
Удаление второй фосфатной группы приводит к выделению эквивалентного количества энергии и образованию аденозинмонофосфата (АМФ):
АДФ + Н20 → АМФ +Н3РО4 + 30,6 кДж.
Примечание:
Конкретные механизмы использования АТФ для совершения всех этих работ связаны с переносом с помощью ферментов киназ концевых фосфатных
групп АТФ на другие молекулы-акцепторы. В результате образуются фосфорилированные продукты, способные активно участвовать в других химических реакциях либо выполнять работу, связанную с
жизнедеятельностью клетки. Например, перенос концевой фосфатной группы от молекулы АТФ на сократительный белок обеспечивает сокращение мышечного волокна.
ВЫВОД АТФ — это универсальный переносчик основной аккумулятор энергии, получаемой клеткой в процессах катаболизма или в ходе
реакций
фото- и хемосинтеза. Накопленная таким образом энергия затем используется клеткой для биосинтеза необходимых ей макромолекул и построения
клеточных структур, а также для обеспечения процессов жизнедеятельности клетки.
Энергетический обмен в клетках большинства аэробных организмов в присутствии кислорода состоит из последовательных этапов:
1.подготовительного,
2.бескислородного (анаэробного),
3. кислородного (аэробного),
в ходе, которых органические вещества расщепляются до совсем простых, бедных энергией неорганических соединений, таких как С02,
Н20 и др. В клетках анаэробных организмов, а также в клетках аэробов при недостатке кислорода осуществляются лишь первые два этапа с образованием не полностью окисленных органических
соединений, еще богатых энергией.
Процесс биологического окисления сходен с горением.
Отличие процессов горения и окисления:
— биологическое окисление происходит
поэтапно:
— энергия выделяется порциями и успевает
запасаться в биологических аккумуляторах
Сущность бескислородного и кислородного этапов энергетического обмена целесообразно рассмотреть на примере биологического окисления глюкозы,
так как главным источником энергии в клетке служат углеводы.
Электроны в составе органических веществ находятся на высоких энергетических уровнях. Перемещаясь на более низкий энергетический уровень
своей или иной молекулы или атома, электроны теряют энергию
Основная энергия глюкозы сосредоточена в ее высокоэнергетических электронах атомов водорода, конечным акцептором которых при аэробном
метаболизме служит молекулярный кислород.
Примечание:
При полном биологическом окислении перенос электронов с глюкозы на кислород осуществляется через ряд промежуточных реакций, сопровождающихся
выделением лишь небольшого количества энергии.
В цепи этих реакций атомы водорода (электроны вместе с протонами), с помощью специальных ферментов − дегидрогеназ, отнимаются от
глюкозы и продуктов ее метаболизма и переносятся на первичные акцепторы и переносчики электронов и водорода никотинамидадениндинуклеотид (НАД) и реже — на флавинадениндинуклеотид
(ФАД).
Этот процесс можно выразить следующим общим уравнением:
АН2 + НАД → А + НАДН2 (молекулярная форма записи)
НАД+ и НАДН + Н+ (ионная форма записи)
Восстановленные молекулы НАДН2 и ФАДН2 служат резервуарами высокоэнергетических электронов, при передаче которых на кислород
освобождается энергия для синтеза АТФ.
Этот процесс осуществляется при участии цепи переноса электронов или дыхательной цепи, встроенной во внутреннюю мембрану митохондрий и включающей
несколько дыхательных ферментов. Благодаря этому, электроны при их последовательном переносе от одного дыхательного фермента к другому и к кислороду освобождают свою энергию не одновременно,
как при обычном химическом взаимодействии водорода и кислорода, а небольшими порциями, которые клетка может использовать для синтеза АТФ.
Таким образом, при полном биологическом окислении большинство электронов глюкозы освобождают заключенную в них энергию, «падая» по цепям
ферментативных реакций в направлении глюкоза → НАДН2 → электрон-транспортная цепь → кислород.
Как видим, конечным акцептором электронов в реакциях расщепления глюкозы служит молекулярный кислород, и в этом заключается его главная роль в
процессах биологического окисления.
Характеристика этапов энергетического обмена
1.
Подготовительный этап.
Белки гидролизуются до аминокислот, жиры — до глицерина и жирных кислот, нуклеиновые кислоты — до нуклеотидов, полисахариды — до моносахаридов.
При этом освобождается незначительное количество энергии, которая рассеивается в виде тепла.
Результатом данного этапа является образование мономеров органических соединений и выделение энергии в виде тепла
2. Бескислородный (анаэробный) этап.
Простые органические соединения, образовавшиеся на подготовительном этапе, подвергаются дальнейшему распаду, который происходит в гиалоплазме
клетки. Сложный процесс ферментативного бескислородного расщепления глюкозы, в результате которого шестиуглеродная глюкоза расщепляется на две трехуглеродные молекулы пировиноградной кислоты,
называется гликолизом (греч. glykys — сладкий). Он осуществляется при участии специальных ферментов, составляющих своеобразный ферментативный конвейер, и не нуждается в присутствии
молекулярного кислорода.
Суммарное уравнение гликолиза имеет следующее выражение:
С6Н1206 + 2НАД +2АДФ + 2ФН → 2СН3СОСООН + 2НАДН2 + 2АТФ
Сравнивая число атомов в двух молекулах пировиноградной кислоты и в молекуле глюкозы, можно видеть, что в процессе гликолиза молекула глюкозы
не только расщепляется на две трехуглеродные молекулы пировиноградной кислоты, но и теряет четыре атома водорода, т. е. происходит ее окисление. Акцептором атомов водорода в этих реакциях, как
это уже отмечалось выше, служат молекулы НАД.
Результатом гликолиза является образование двух молекул пировиноградной кислоты, 2 АТФ и восстановленного 2
НАДН2.
В молекулах АТФ аккумулируется более 31% высвобождающейся при этом энергии. Образование АТФ в процессе гликолиза происходит посредством
субстратного фосфорилирования, при котором фермент переносит фосфатную группу непосредственно от субстрата на АДФ.
Дальнейшие превращения пировиноградной кислоты, соли которой называются пируваты, зависят от условий среды и вида клеток.
Пути метаболизма ПВК
При отсутствии кислорода у многих бактерий, грибов и простейших пируват в присутствии НАДН2, образовавшегося в процессе гликолиза,
восстанавливается до молочной кислоты, соли которой называются лактаты. Этот процесс называется молочнокислым брожением. Молочная кислота может также образовываться в мышечных клетках человека
при недостатке кислорода, например, при тяжелой физической работе.
Уравнение реакции молочнокислого брожения (бактерии, грибы) имеет следующее выражение:
СН3СОСООН + НАДН2 → СН3СНОНСООН + НАД.
В дрожжах и во многих растительных клетках в анаэробных условиях пируват расщепляется до этанола и С02. Спиртовое
брожжение:
СН3СОСООН + НАДН2 → СН3СН2ОН + С02 + НАД.
3. В аэробных условиях наступает третий — кислородный этап энергетического обмена, в ходе которого продукты гликолиза в цепи
последовательных реакций расщепляются до углекислого газа и воды. Этот заключительный период энергетического обмена называется клеточным дыханием, которое следует отличать от внешнего
дыхания, обеспечивающего поглощение кислорода из окружающей среды и выделение в нее С02.
Клеточное дыхание включает три группы реакций:
1. Образование ацетилкофермента А.
2. Цикл трикарбоновых кислот.
3. Перенос электронов по дыхательной цепи и окислительное фосфорилирование.
Первые две группы реакций происходят в матриксе митохондрий, а третья — на их внутренней мембране.
1. Ацетилкофермент А (ацетил-КоА) образуется при окислительном декарбоксилировании пирувата с участием целой группы ферментов. В результате
ряда реакций ацетильная группа пирувата (СН3СО—) присоединяется к переносчику этих групп — сложному соединению коферменту A (HS-KoA), образуя весьма активное и богатое энергией соединение — ацетил-КоА, служащее «топливом», поступающим в цикл
трикарбоновых кислот. Окисление молекулы пирувата сопровождается восстановлением молекулы НАД до НАДН2 и образованием молекулы диоксида углерода по уравнению:
СН3СОСООН + НАД + КоА → Ацетил-КоА + НАДН2 + С02.
Поскольку в результате окисления одной молекулы глюкозы образуются две молекулы пирувата, число молекул всех компонентов реакции необходимо
удвоить.
2. Цикл трикарбоновых кислот (цикл лимонной кислоты) получил это название вследствие участия в нем лимонной кислоты (имеющей три
карбоксильные группы), которая является первым его продуктом. Этот цикл называют также циклом Кребса — по фамилии английского ученого, который открыл его в 1937 г. Цикл Кребса состоит из
восьми последовательных реакций, образующих замкнутый круг.
В начале цикла ацетильная группа, освобождающаяся при гидролизе ацетил-КоА, соединяется с четырехуглеродной щавелевоуксусной кислотой
(оксалоацетатом) и образует шестиуглеродную лимонную кислоту. По ходу работы цикла два из шести атомов углерода окисляются до С02 и вновь образуется одна молекула оксалоацетата,
которая готова начать новый оборот цикла. Энергия, выделяемая при окислении углеродных атомов, используется для образования одной молекулы АТФ, трех молекул НАДН2, одной молекулы
ФАДН2.
Итоговое уравнение окислительно-восстановительных реакций в цикле Кребса имеет следующее выражение:
Оксалоацетат + Ацетил-КоА+АДФ +ФН
ЗНАД + ФАД → Оксалоацетат + 2С02 + КоА + АТФ + ЗНАДН2 + ФАДН2.
Образующаяся в цикле Кребса углекислота высвобождается из митохондрии путем диффузии и выводится из клетки.
При окислении одной молекулы глюкозы совершается два оборота цикла Кребса – по одному на каждую из двух образующихся при этом молекул
ацетил-КоА.
Таким образом, в результате реакций гликолиза и цикла Кребса молекула глюкозы расщепляется до диоксида углерода, который выделяется из клетки, а
высвободившаяся при этом энергия частично расходуется на синтез АТФ, но, в основном, аккумулируется в нагруженных высокоэнергетическими электронами восстановленных молекулах НАДН2 и
ФАДН2.
3. Конечная стадия клеточного дыхания включает перенос электронов от восстановленных молекул НАДН2 и ФАДН2 по цепи
транспорта электронов, состоящей из девяти ферментов, встроенных во внутреннюю мембрану митохондрий, на молекулу кислорода.
Энергия, выделяющаяся небольшими порциями при переносе электронов, используется для синтеза АТФ из АДФ. Этот процесс называется
окислительным фосфорилированием.
Для объяснения механизма преобразования энергии при окислительном фосфорилировании английский ученый П. Митчел предложил хемиосмотическую
теорию, ставшую ныне общепризнанной.
Согласно этой теории, восстановленные молекулы НАДН2 и ФАДН2 передают атомы водорода в дыхательную цепь, содержащуюся во
внутренней мембране митохондрий, где от них отщепляются электроны (ē) с образованием протонов (Н+). Электроны переносятся по дыхательной цепи на молекулу кислорода и активируют ее.
Активированный кислород сразу же реагирует с образовавшимися протонами, в результате чего выделяется вода (на каждые две молекулы НАДН2 восстанавливается одна молекула кислорода
выделяются две молекулы воды).
В процессе переноса электронов по электрон-транспортной цепи они освобождают свою энергию, которая используется некоторыми ферментами дыхательной
цепи, работающими как протонные насосы для перекачивания ионов водорода (Н+) из матрикса митохондрий в межмембранное пространство. Здесь происходит их накопление, так как внутренняя
мембрана митохондрий непроницаема для ионов (образуется Н+- резервуар).
В матриксе, на внутренней стороне мембраны, концентрация протонов уменьшается, а ионы гидроксила (ОН-) оказываются в избытке.
В результате наружная сторона внутренней мембраны митохондри?