Какой пигмент содержится в хлоропластах

Хлоропласты – двухмембранные органоиды растительных клеток, именно они играют ключевую роль в одном из самых важных биологических процессов в природе – фотосинтезе. В частности именно хлоропласты в процессе фотосинтеза выделяют зеленый пигмент хлорофилл, благодаря которому листья деревьев приобретают зеленый цвет (впрочем, не только листья, но и многие другие представители растительного мира, например водоросли). Какое строение хлоропластов, какие функции и процессы они осуществляются в жизнедеятельности клетки, об этом читайте далее.
Количество хлоропластов в растительной клетке может быть разным, у некоторых водорослей в клетке содержится лишь один большой хлоропласт, часто причудливой формы, в то время как в клетках некоторых высших растений находится множество хлоропластов. Особенно их много в так званных мезофильных тканях листьев, там одна клетка может иметь в себе до сотни хлоропластов.
Строение хлоропластов
Устройство хлоропласта включает в себя внутреннюю и внешнюю мембрану, (как и в клетке, они играют роль защитного барьера), межмембранное пространство, строму, тилакоиды, граны, ламеллы, люмен.
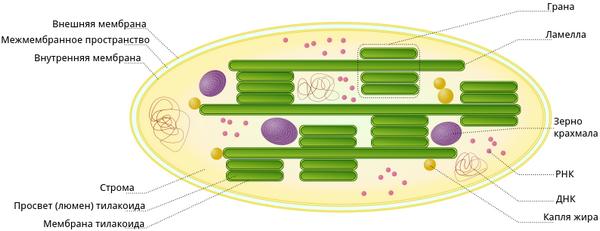
Вот так строение хлоропласта выглядит на картинке.
Как видим с картинки внутри хлоропласта имеется полужидкое пространство, именуемое стромой и приплюснутые диски – это тилакоиды. Последние объединены в стопки, названные гранамы, и сами граны соединены друг с другом при помощи длинных тилакоид, которые называют ламеллами. Именно в тилакоидах находится важный зеленый пигмент – хлорофилл.
В полужидкой строме хлоропласта находятся его молекулы ДНК и РНК, а также рибосомы, обеспечивающие этому важному органоиду некую автономность внутри клетки. Помимо этого в строме хлоропласта есть зерна крахмала, которые образуются при избытке углеводов, образованных при фотосинтетической активности.
Функции хлоропластов
Самая важная функция хлоропласта – это, конечно же, осуществление фотосинтеза. Об этом удивительном процессе на нашем сайте есть отдельная большая статья. Тем не менее, напомним, что при фотосинтезе хлоропластами растительных клеток при помощи солнечного света осуществляется синтез глюкозы из углекислого газа и воды. При этом в качестве важного «побочного продукта» выделяется кислород.
Основным фотосинтезирующим пигментом в этом процессе является хлорофилл, локализированный в мембранах тилакоидов, именно здесь проходят световые реакции фотосинтеза. Кроме хлорофилла тут же присутствуют ферменты и переносчики электронов.
Интересный факт: хлоропласты стараются расположиться в клетке таким образом, чтобы их тилакоидные мембраны находились под прямым углом к солнечному свету. Или говоря простым языком, хлоропласты в клетке всегда тянутся на свет.
Строение хлорофилла
Что же касается строения самого хлорофилла, то он состоит из длинного углеводного хвоста и порфириновой головки. Хвост его гидрофобен, то есть боится влаги, поэтому погружен в тилакоид, головка наоборот любит влагу и находится в жидкой субстанции хлоропласта – строме. Поглощение солнечного света осуществляется именно головкой хлорофилла.
К слову биологами различается несколько разных видов хлорофилла: хлорофилл a, хлорофилл b, хлорофилл c1, хлорофилл c2 и так далее, все они обладают разным спектром поглощения солнечного света. Но больше всего в растениях именно хлорофилла а.
Рекомендованная литература и полезные ссылки
- Белякова Г. А. Водоросли и грибы // Ботаника: в 4 т. / Белякова Г. А., Дьяков Ю. Т., Тарасов К. Л. — М.: Издательский центр «Академия», 2006. — Т. 1. — 320 с. — 3000 экз. — ISBN 5-7695-2731-5.
- Карпов С.А. Строение клетки протистов. — СПб.: ТЕССА, 2001. — 384 с. — 1000 экз. — ISBN 5-94086-010-9.
- Lee, R. E. Phycology, 4th edition. — Cambridge: Cambridge University Press, 2008. — 547 с. — ISBN 9780521682770.
Хлоропласты, видео
И в завершение образовательное видео по теме нашей статьи.
Автор: Павел Чайка, главный редактор журнала Познавайка
При написании статьи старался сделать ее максимально интересной, полезной и качественной. Буду благодарен за любую обратную связь и конструктивную критику в виде комментариев к статье. Также Ваше пожелание/вопрос/предложение можете написать на мою почту pavelchaika1983@gmail.com или в Фейсбук, с уважением автор.
Фотосинтез
связан с избирательным поглощением
пигментами света в видимой части
солнечного спектра. Фотосинтетические
питменты составляют 10-15 % сухой массы
хлоропластов. Они характеризуются
большим разнообразием и по химической
природе делятся на две группы — хлорофиллы
и каротиноиды.
Хлорофиллы
Состав
и структура.
В настоящее время известно несколько
различньк форм хлорофилла, которые
обозначают латинскими буквами. хлоропласты
высших растений содержат хлорофилл а
и хлорофилл b.
Они были идентифицированы русским
ученым М.
С. Цветом
(1906) с помощью разработанного им метода
хроматографии. Структурная формула
хлорофилла, предложенная Г.
Фишером
(1939), получила окончательное подтверждение
в 1960 г. в результате двух независимо
проведенных работ в США и ФРГ по
искусственному синтезу хлорофилла а.
Хлорофилл
— сложный эфир дикарбоновой кислоты
хлорофиллина. у которой одна карбоксильная
группа этерифицирована спиртом фитолом,
а вторая — спиртом метанолом.
Каратиноиды
Световая фаза фотосинтеза
Сущность
световой фазы фотосинтеза состоит в
поглощении лучистой энергии и ее
трансформации в ассимиляционную силу
(АТФ и НАДФН),
необходимую для восстановления углерода
в темповых реакциях. Сложность процессов
преобразования световой энергии в
химическую требует их строгой мембранной
организации. Световая фаза фотосинтеза
происходит в
гранах
хлоропласта.
В
состав фотохимически активных мембран
хлоропластов входят 5
согласованно функционирующих
многокомпонентных белковых комплексов:
светособирающий
комплекс (ССК),ФС
1,ФС
2,цитохромный
комплекс (цитохромы b6
и f),
участвующий в транспорте электронов,
иАТФ-азный
комплекс, обеспечивающий синтез
АТФ.
Организация и функционирование пигментных систем
Пигменты
хлоропластов объединены в функциональные
комплексы — пигментные
системы,
в которых реакционный
центр (РЦ)
хлорофилл
а,
осуществляющий фотосенсибилизацию,
связан процессами переноса энергии с
антенной,
состоящей из светособирающих
пигментов.
Современная
схема фотосинтеза высших растений
включает 2 фотохимические реакции,
осуществляемые с участием 2 различных
ФС. Предположение об их существовании
было высказано Р. Эмерсоном
в 1957 г. на основании обнаруженного им
эффекта усиления действия длинноволнового
(ДВ) красного света (700 нм) совместным
освещением более коротковолновыми
лучами (650 нм).
Впоследствии
с помощью обработки ультразвуком и
поверхностно-активными веществами
и использования дифференциального
центрифугирования в градиенте плотности
сахарозы удалось получить легкие
фрагменты хлоропластов, обогащенные
фракцией ФС1, и более тяжелые фрагменты,
содержащие фракцию ФС2.
Установлено,
что ФС II
поглощает
более коротковолновые лучи
по сравнению с ФС 1.
Фотосинтез идет эффективно только при
их совместном функционировании,
что объясняет эффект усиления Эмерсона.
ФС I:
в качестве РЦ
выступает димер хлорофилла
а
с максимумом поглощения света 700
нм
(P700),
роль антенного
компонента
играют также хлорофиллы
а675-695.
Первичным
акцептором
электронов в этой системе является
мономерная форма
хлорофилла а695,
вторичными
акцепторами
— железосерные белки (-FeS).
Комплекс
ФС 1
под действием света восстанавливает
железосодержащий белок — ферредоксин
(Фд)
и окисляет
медьсодержащий белок- пластоцианин
(Пц).
ФC II:
включает РЦ,
содержащий хлорофилл
а (P680)
и антенные
пигменты —
хлорофиллы
a670-683.
Первичным
акцептором
электронов является феофитин
(фф),
передающий электроны на пластохинон.
В состав ФС II входит также белковый
комплекс S-системы,
окисляющий
воду,
и переносчик
электронов
Z.
Этот комплекс функционирует с участием
Mn,
Cl
и Mg.
ФС
II восстанавливает
пластохинон
(PQ)
и окисляет
воду
с выделением О2
и протонов.
Связующим
звеном между ФС II и ФС 1
служат фонд пластохинонов,
белковый цитохромный
комплекс
(b6,
f)
и пластоцианин.
На
один
РЦ
приходится примерно 300
молекул пигментов,
которые входят в
состав антенных или светособирающих
комплексов (ССК).
Светособирающий
белковый комплекс
ламелл связан с ФСII и содержит
хлорофиллы
а, b
и каротиноиды.
В каждом ССК содержится от
120 до 240
молекул хлорофилла. Антенные
комплексы
входят в состав ФС1 и ФСII
(фокусирующие антенные компоненты ФС).
Антенный
белковый комплекс
ФС 1
содержит 110
молекул
хлорофиллов а680-695
на один P700,
из них 60 молекул — компоненты антенного
комплекса, который можно рассматривать
как ССК ФС1. Антенный комплекс ФС1 также
содержит -каротин.
Антенный
белковый комплекс
ФС II
содержит 40 молекул хлорофиллов а670-683
на один P680
и
-каротин.
Хромопротеины
антенных комплексов не обладают
фотохимической активностью. Их роль
состоит в поглощении и передаче энергии
квантов на небольшое количество молекул
РЦ
P700
и P680,
каждая из которых связана с цепью
транспорта электронов и осуществляет
фотохимическую реакцию. Организация
электронно-транспортных
цепей (ЭТЦ)
при всех молекулах хлорофилла
нерациональна, так как даже на прямом
солнечном свету на молекулу пигмента
кванты света попадают не
чаще одного раза за 0,1 с.
Передача
энергии в светособирающих комплексах
и ФС происходит путем
индуктивного резонанса.
Условиями для резонансного переноса
энергии служат сближенное
расположение
и перекрытие частот
осциллирующих
электронных систем
у двух взаимодействующих молекул. Причем
миграция энергии осуществляется от
коротковолновых пигментов к более
длинноволновым.
Квант света мигрирует в ФС, пока не
попадет в стоковую
молекулу (ловушку)
— пигмент
реакционного центра,
который является самым длинноволновым
и поэтому неспособным возбуждать
соседние молекулы. Энергия
возбужденного пигмента РЦ расходуется
на фотохимическую работу.
Квант света обычно передается на стоковую
молекулу через
несколько сотен передач,
каждая передача осуществляется за
10-12
с,
100
передач-соответственно за 10-10
с.
Экспериментально установлено, что 90 %
энергии возбуждения хлорофилла а
может быть передано на P700
менее чем за
10-9
с.
Соседние файлы в папке Физиология и биохимия
- #
- #
- #
- #
- #
- #
- #
Ð Ð°Ð·Ð¼ÐµÑ Ñ Ð»Ð¾ÑоплаÑÑов колеблеÑÑÑ Ð¾Ñ 4 до 10 мкм. ЧиÑло Ñ Ð»Ð¾ÑоплаÑÑов обÑÑно ÑоÑÑавлÑÐµÑ Ð¾Ñ 20 до 100 на клеÑкÑ. ХимиÑеÑкий ÑоÑÑав Ñ Ð»Ð¾ÑоплаÑÑов доÑÑаÑоÑно Ñложен и Ð¼Ð¾Ð¶ÐµÑ Ð±ÑÑÑ Ð¾Ñ Ð°ÑакÑеÑизован ÑледÑÑÑими ÑÑедними даннÑми (% на ÑÑÑ ÑÑ Ð¼Ð°ÑÑÑ): белок â 35â55; Ð»Ð¸Ð¿Ð¸Ð´Ñ â 20â30; ÑÐ³Ð»ÐµÐ²Ð¾Ð´Ñ â 10; Ð ÐÐ â 2â3; ÐÐÐ â до 0,5; Ñ Ð»Ð¾ÑоÑилл â 9; каÑоÑÐ¸Ð½Ð¾Ð¸Ð´Ñ â 4,5. Ðажно оÑмеÑиÑÑ, ÑÑо многие белки Ñ Ð»Ð¾ÑоплаÑÑов обладаÑÑ ÑеÑменÑаÑивной акÑивноÑÑÑÑ. ÐейÑÑвиÑелÑно, в Ñ Ð»Ð¾ÑоплаÑÑÐ°Ñ ÑоÑÑедоÑоÑÐµÐ½Ñ Ð²Ñе ÑеÑменÑÑ, пÑинимаÑÑие ÑÑаÑÑие в пÑоÑеÑÑе ÑоÑоÑинÑеза (окиÑлиÑелÑно-воÑÑÑановиÑелÑнÑе, ÑинÑеÑÐ°Ð·Ñ Ð³Ð¸Ð´ÑолазÑ). РнаÑÑоÑÑее вÑÐµÐ¼Ñ Ð´Ð¾ÐºÐ°Ð·Ð°Ð½Ð¾, ÑÑо в Ñ Ð»Ð¾ÑоплаÑÑÐ°Ñ , Ñак же как и в миÑÐ¾Ñ Ð¾Ð½Ð´ÑиÑÑ , имееÑÑÑ ÑÐ²Ð¾Ñ Ð±ÐµÐ»Ð¾ÐºÑинÑезиÑÑÑÑÐ°Ñ ÑиÑÑема. Ðногие из ÑеÑменÑов, локализованнÑÑ Ð² Ñ Ð»Ð¾ÑоплаÑÑÐ°Ñ , ÑвлÑÑÑÑÑ Ð´Ð²ÑÑ ÐºÐ¾Ð¼Ð¿Ð¾Ð½ÐµÐ½ÑнÑми. Ðо Ð¼Ð½Ð¾Ð³Ð¸Ñ ÑлÑÑаÑÑ Ð¿ÑоÑÑеÑиÑеÑÐºÐ°Ñ Ð³ÑÑппа ÑеÑменÑов â ÑÑо ÑазлиÑнÑе виÑаминÑ. Ð Ñ Ð»Ð¾ÑоплаÑÑÐ°Ñ ÑоÑÑедоÑоÑÐµÐ½Ñ Ð¼Ð½Ð¾Ð³Ð¸Ðµ виÑÐ°Ð¼Ð¸Ð½Ñ Ð¸ Ð¸Ñ Ð¿ÑоизводнÑе (виÑÐ°Ð¼Ð¸Ð½Ñ Ð³ÑÑÐ¿Ð¿Ñ Ð, Ð, Ð, D). Ð Ñ Ð»Ð¾ÑоплаÑÑÐ°Ñ Ð½Ð°Ñ Ð¾Ð´Ð¸ÑÑÑ 80% Fe, 70% Zn, около 50% Си Ð¾Ñ Ð²Ñего колиÑеÑÑва ÑÑÐ¸Ñ ÑлеменÑов в лиÑÑе.
ÐнÑÑÑеннее ÑÑÑоение Ñ Ð»Ð¾ÑоплаÑÑов, Ð¸Ñ ÑлÑÑÑаÑÑÑÑкÑÑÑа бÑли ÑаÑкÑÑÑÑ Ñ Ð¸ÑполÑзованием ÑлекÑÑонного микÑоÑкопа. ÐказалоÑÑ, ÑÑо Ñ Ð»Ð¾ÑоплаÑÑÑ Ð¾ÐºÑÑÐ¶ÐµÐ½Ñ Ð´Ð²Ð¾Ð¹Ð½Ð¾Ð¹ мембÑаной. ТолÑина каждой мембÑÐ°Ð½Ñ 7,5â10 нм, ÑаÑÑÑоÑние Ð¼ÐµÐ¶Ð´Ñ Ð½Ð¸Ð¼Ð¸ 10â30 нм. ÐнÑÑÑеннее пÑоÑÑÑанÑÑво Ñ Ð»Ð¾ÑоплаÑÑов заполнено беÑÑвеÑнÑм ÑодеÑжимÑм â ÑÑÑомой и пÑонизано мембÑанами (ламеллами). ÐамеллÑ, ÑоединеннÑе дÑÑг Ñ Ð´ÑÑгом, обÑазÑÑÑ ÐºÐ°Ðº Ð±Ñ Ð¿ÑзÑÑÑкиâÑÐ¸Ð»Ð°ÐºÐ¾Ð¸Ð´Ñ (гÑеÑ. «ÑилакоидеÑ» â меÑковиднÑй). Ð Ñ Ð»Ð¾ÑоплаÑÑÐ°Ñ ÑодеÑжаÑÑÑ ÑÐ¸Ð»Ð°ÐºÐ¾Ð¸Ð´Ñ Ð´Ð²ÑÑ Ñипов. ÐоÑоÑкие ÑÐ¸Ð»Ð°ÐºÐ¾Ð¸Ð´Ñ ÑобÑÐ°Ð½Ñ Ð² паÑки и ÑаÑÐ¿Ð¾Ð»Ð¾Ð¶ÐµÐ½Ñ Ð´ÑÑг над дÑÑгом, Ð½Ð°Ð¿Ð¾Ð¼Ð¸Ð½Ð°Ñ ÑÑÐ¾Ð¿ÐºÑ Ð¼Ð¾Ð½ÐµÑ. ÐÑи ÑÑопки назÑваÑÑÑÑ Ð³Ñанами, а ÑоÑÑавлÑÑÑие Ð¸Ñ ÑÐ¸Ð»Ð°ÐºÐ¾Ð¸Ð´Ñ â Ñилакоидами гÑан. ÐÐµÐ¶Ð´Ñ Ð³Ñанами паÑаллелÑно дÑÑг дÑÑÐ³Ñ ÑаÑполагаÑÑÑÑ Ð´Ð»Ð¸Ð½Ð½Ñе ÑилакоидÑ. СоÑÑавлÑÑÑие Ð¸Ñ Ð»Ð°Ð¼ÐµÐ»Ð»Ñ Ð¿Ð¾Ð»ÑÑили название â ÑÐ¸Ð»Ð°ÐºÐ¾Ð¸Ð´Ñ ÑÑÑомÑ. ÐÐµÐ¶Ð´Ñ Ð¾ÑделÑнÑми Ñилакоидами в ÑÑÐ¾Ð¿ÐºÐ°Ñ Ð³Ñан имеÑÑÑÑ Ñзкие Ñели. ТилакоиднÑе мембÑÐ°Ð½Ñ ÑодеÑÐ¶Ð°Ñ Ð±Ð¾Ð»ÑÑое колиÑеÑÑво белков, ÑÑаÑÑвÑÑÑÐ¸Ñ Ð² ÑоÑоÑинÑезе. Ð ÑоÑÑаве инÑегÑалÑнÑÑ Ð¼ÐµÐ¼Ð±ÑаннÑÑ Ð±ÐµÐ»ÐºÐ¾Ð² имееÑÑÑ Ð¼Ð½Ð¾Ð³Ð¾ гидÑоÑобнÑÑ Ð°Ð¼Ð¸Ð½Ð¾ÐºÐ¸ÑлоÑ. ÐÑо ÑÐ¾Ð·Ð´Ð°ÐµÑ Ð±ÐµÐ·Ð²Ð¾Ð´Ð½ÑÑ ÑÑÐµÐ´Ñ Ð¸ Ð´ÐµÐ»Ð°ÐµÑ Ð¼ÐµÐ¼Ð±ÑÐ°Ð½Ñ ÑÑабилÑнее. Ðногие белки ÑилакоиднÑÑ Ð¼ÐµÐ¼Ð±Ñан поÑÑÑÐ¾ÐµÐ½Ñ Ð² виде векÑоÑов и гÑаниÑÐ°Ñ Ñ Ð¾Ð´Ð½Ð¾Ð¹ ÑÑоÑÐ¾Ð½Ñ Ñо ÑÑÑомой, а Ñ Ð´ÑÑгой конÑакÑиÑÑÑÑ Ñ Ð²Ð½ÑÑÑенним пÑоÑÑÑанÑÑвом Ñилакоида.
ÐÑноÑиÑелÑно ÑвÑзи Ð¼ÐµÐ¶Ð´Ñ Ð»Ð°Ð¼ÐµÐ»Ð»Ð°Ð¼Ð¸ гÑан и ламеллами ÑÑÑÐ¾Ð¼Ñ Ð¸Ð¼ÐµÑÑÑÑ ÑазнÑе ÑоÑки зÑениÑ. Т. ÐÐµÐ¹ÐµÑ Ð¿Ñедложил гÑанÑлÑÑно-ÑеÑеÑÑаÑÑÑ Ð¼Ð¾Ð´ÐµÐ»Ñ, ÑоглаÑно коÑоÑой внÑÑÑенние пÑоÑÑÑанÑÑва вÑÐµÑ Ñилакоидов ÑÐ¾ÐµÐ´Ð¸Ð½ÐµÐ½Ñ Ð¼ÐµÐ¶Ð´Ñ Ñобой. Таким обÑазом, в Ñ Ð»Ð¾ÑоплаÑÑÐ°Ñ Ð¸Ð¼ÐµÐµÑÑÑ ÐºÐ°Ðº Ð±Ñ Ð´Ð²Ð° ÑазделÑнÑÑ Ð¿ÑоÑÑÑанÑÑва â внÑÑÑеннее (внÑÑÑи Ñилакоидов) и внеÑнее (вне Ñилакоидов). У болÑÑинÑÑва водоÑоÑлей гÑан неÑ, а Ð»Ð°Ð¼ÐµÐ»Ð»Ñ ÑобÑÐ°Ð½Ñ Ð² гÑÑÐ¿Ð¿Ñ (паÑки) по 2â8 ÑÑÑк. Ðе во вÑÐµÑ ÑлÑÑаÑÑ Ð¸ Ñ Ð²ÑÑÑÐ¸Ñ ÑаÑÑений Ñ Ð»Ð¾ÑоплаÑÑÑ Ð¸Ð¼ÐµÑÑ Ð³ÑаналÑнÑÑ ÑÑÑÑкÑÑÑÑ. Так, в лиÑÑÑÑÑ ÐºÑкÑÑÑÐ·Ñ Ð¸Ð¼ÐµÑÑÑÑ Ð´Ð²Ð° вида Ñ Ð»Ð¾ÑоплаÑÑов. РклеÑÐºÐ°Ñ Ð¼ÐµÐ·Ð¾Ñилла ÑодеÑжаÑÑÑ Ð¼ÐµÐ»ÐºÐ¸Ðµ Ñ Ð»Ð¾ÑоплаÑÑÑ Ð³ÑаналÑного ÑÑÑоениÑ. РклеÑÐºÐ°Ñ Ð¾Ð±ÐºÐ»Ð°Ð´ÐºÐ¸, окÑÑжаÑÑÐ¸Ñ Ð»Ð¸ÑÑовÑе ÑоÑÑдиÑÑÑе пÑÑки, Ñ Ð»Ð¾ÑоплаÑÑÑ ÐºÑÑпнÑе и гÑан не ÑодеÑжаÑ. Ð ÑÑÑоме Ñ Ð»Ð¾ÑоплаÑÑов Ð½Ð°Ñ Ð¾Ð´ÑÑÑÑ Ð½Ð¸Ñи ÐÐÐ, ÑибоÑомÑ, кÑÐ°Ñ Ð¼Ð°Ð»ÑнÑе зеÑна.

Ð¡Ñ ÐµÐ¼Ð° гÑанÑлÑÑно-ÑеÑÑаÑой ÑÑÑÑкÑÑÑÑ Ñ Ð»Ð¾ÑоплаÑÑов.
ÐÑновной ÑеÑменÑ, обеÑпеÑиваÑÑий ÑÑвоение ÑглекиÑлого газа, â ÑибÑзолобиÑоÑÑаÑкаÑбокÑилазаокÑигеназа (ÑокÑаÑенно Ð ÐФ â каÑбокÑилазаокÑигеназа) Ñакже ÑаÑположен в ÑÑÑоме. ÐеленÑй Ð¿Ð¸Ð³Ð¼ÐµÐ½Ñ Ñ Ð»Ð¾ÑоÑилл в виде комплекÑа Ñ Ð±ÐµÐ»ÐºÐ°Ð¼Ð¸ (пигменÑ-белковÑе комплекÑÑ) ÑоÑÑедоÑоÑен главнÑм обÑазом в ÑÐ¸Ð»Ð°ÐºÐ¾Ð¸Ð´Ð°Ñ Ð³Ñан и ÑаÑÑиÑно в ÑÐ¸Ð»Ð°ÐºÐ¾Ð¸Ð´Ð°Ñ ÑÑÑомÑ. РмембÑÐ°Ð½Ð°Ñ Ñилакоидов гÑан локализован ÑеÑменÑ, каÑализиÑÑÑÑий ÑинÑез ÐТФ (ÐТФ-ÑинÑаза). ÐÑÐ¾Ñ ÑеÑÐ¼ÐµÐ½Ñ ÑвÑзан Ñ Ð±ÐµÐ»ÐºÐ¾Ð¼, ÑаÑположеннÑм в Ñамой мембÑане.

В обзорной статье В.Г.Ладыгина и Г.Н.Ширшиковой изложены современные представления о функциях каротиноидов — желтых, красных и оранжевых пигментов — у растений. Каротиноиды играют очень важную роль в работе молекулярной машины фотосинтеза. Они выполняют три основные функции: фотозащитную (защищают хлорофилл и другие уязвимые компоненты фотосистем от светового «перевозбуждения»), светособирающую (что позволяет растениям использовать энергию света в синей области спектра — задача, с которой хлорофилл не может справиться без помощи каротиноидов) и структурную (служат необходимыми структурными элементами, «кирпчиками» фотосистем).
Каротиноиды — широко распространенный класс пигментов, встречающийся у бактерий, одноклеточных эукариот, грибов, растений и животных. В отличие от ряда других пигментов, таких как гем (окрашивающий кровь и мышцы млекопитающих в красный цвет) или хлорофилл (ответственный за зеленую окраску растений), молекулы каротиноидов не содержат металлов. Они состоят только из углерода, водорода и кислорода, и их способность «работать» с квантами света определяется системой сопряженных двойных связей между атомами углерода, выстроенными в цепочку. Сопряженными называются двойные связи, разделенные одной простой связью.
Каротиноиды поглощают свет с длиной волны 280–550 нм (это зеленая, синяя, фиолетовая, ультрафиолетовая области спектра). Чем больше в молекуле сопряженных двойных связей, тем больше длина волны поглощаемого света. Соответственно меняется и окраска пигмента. Каротиноиды, имеющие 3–5 сопряженных двойных связей, бесцветны, они поглощают свет в ультрафиолетовой области. Дзета-каротин с семью связями имеет желтую окраску, нейроспорин с девятью связями — оранженвую, ликопин с 11 связями — оранжево-красную.

Функции каротиноидов в живой природе не ограничиваются работой со светом, порой они играют важную роль в обмене веществ (вспомним, например, витамин А — производное бета-каротина). И все же главные их функции (будь то в органах зрения животных или в хлоропластах — органеллах фотосинтеза растений) неразрывно связаны со светом. В статье Ладыгина и Ширшиковой рассматривается роль каротиноидов в хлоропластах — органеллах растительной клетки, которые ведут свое происхождение от симбиотических цианобактерий. Основная функция хлоропластов — фотосинтез, то есть производство органики из углекислого газа за счет энергии солнечного света. В мембранах хлоропластов расположены белково-пигментные комплексы — фотосистемы I и II, в состав которых входят разнообразные белки, а также пигменты — хлорофиллы и каротиноиды.
Хлорофилл — основной фотосинтетический пигмент — сам по себе способен поглощать и использовать свет только в красной области спектра (650–710 нм). Каротиноиды поглощают сине-зеленый свет и передают его энергию хлорофиллам. Эта функция каротиноидов — светособирающая — особенно важна для водорослей, поскольку сине-зеленый свет проникает в толщу воды гораздо глубже, чем красный.
Вторая функция каротиноидов в хлоропластах — светозащитная. Они защищают фотосистемы от световых «перегрузок», которые могут приводить к сверхвозбуждению и сбоям в работе фотосистем. Каротиноиды служат своего рода «аварийными клапанами», позволяющими сбросить избыточную энергию, перевести ее в тепло. Каротиноиды справляются с этой задачей несколькими разными способами: просто «фильтруя» поступающий свет, забирая на себя избыточную световую энергию, или снимая энергию с перевозбужденного хлорофилла. Каротиноиды могут также «тушить» активные формы кислорода, то есть служат антиоксидантами.
Одним из способов, при помощи которых каротиноиды «сбрасывают» лишнюю энергию при избыточном освещении, являются циклические химические реакции, в ходе которых одни каротиноиды превращаются в другие. Самая распространенная из этих реакций получила название виолаксантинового цикла. На сильном свету каротиноид виолаксантин превращается в зеаксантин, при этом выделяется кислород. Когда освещенность снижается, зеаксантин превращается обратно в виолаксантин, при этом кислород поглощается. Обе реакции — и прямая, и обратная — катализируются ферментами, гены которых расположены в хромосоме хлоропласта, а не в центральном (ядерном) геноме растительной клетки.
Третья функция каротиноидов — структурная. Каротиноиды — обязательные структурные компоненты фотосинтетических мембран хлоропластов. Экспериментально показано, что без каротиноидов фотосистемы становятся нестабильными. Молекулы каротиноидов занимают строго определенные положения в фотосистемах, и без них вся конструкция попросту разваливается.
Авторы отмечают, что в последние годы о каротиноидах стало известно много нового, однако целый ряд подробностей еще предстоит выяснить. В частности, не до конца еще понятно эволюционное происхождение каротиноидов, а также биохимических и фотохимических реакций с их участием. Неясно, в какой степени можно использовать каротиноиды в филогенетике, то есть для реконструкции путей эволюционного развития организмов. Во многих старых работах наборы каротиноидов, характерные для той или иной группы организмов, использовались как важный таксономический признак. Не совсем ясно, насколько такие признаки надежны, особенно если учесть, что одни и те же каротиноиды можно встретить, например, в хлоропластах растений и в глазах млекопитающих.