Какое азотистое основание не содержится в днк
Нуклеиновые кислоты — это
природные высокомолекулярные соединения (полинуклеотиды), которые играют
огромную роль в хранении и передаче наследственной информации в живых
организмах.
Молекулярная масса нуклеиновых кислот
может меняться от сотен тысяч до десятков миллиардов. Они были открыты и
выделены из клеточных ядер еще в XIX в., однако их биологическая роль была
выяснена только во второй половине XX в.
В состав
нуклеотида — структурного звена нуклеиновых кислот — входят три составные
части:
1) азотистое основание — пиримидиновое или
пуриновое
Пиримидиновые
основания
– производные пиримидина, входящие в состав нуклеиновых кислот: урацил,
тимин, цитозин.
Для оснований, содержащих группу –ОН, характерно подвижное равновесие
структурных изомеров, обусловленное переносом протона от кислорода к азоту и
наоборот:
Пуриновые
основания
— производные пурина, входящие в состав нуклеиновых кислот: аденин, гуанин.
Гуанин
существует в виде двух структурных изомеров:
2) моносахарид
Рибоза и 2-дезоксирибоза относятся к моносахаридам, содержащим пять углеродных
атомов. В состав нуклеиновых кислот они входят в циклических β-формах:
3) остаток фосфорной кислоты
ДНК и
РНК
В
зависимости от того, какой моносахарид содержится в структурном звене
полинуклеотида — рибоза или 2-дезоксирибоза, различают
·
рибонуклеиновые кислоты (РНК) и
·
дезоксирибонуклеиновые кислоты (ДНК)
В
главную (сахарофосфатную) цепь РНК входят остатки рибозы, а в ДНК – 2-дезоксирибозы.
Нуклеотидные звенья макромолекул ДНК могут содержать аденин, гуанин, цитозин
и тимин. Состав РНК отличается тем, что вместо тимина присутствует
урацил.
Молекулярная
масса ДНК достигает десятков миллионов а.е.м. Это самые длинные из известных
макромолекул. Значительно меньше молекулярная масса РНК (от нескольких сотен до
десятков тысяч). ДНК содержатся в основном в ядрах клеток, РНК – в рибосомах и
протоплазме клеток.
При
описании строения нуклеиновых кислот учитывают различные уровни организации
макромолекул: первичную и вторичную структуру.
·
Первичная структура нуклеиновых
кислот – это нуклеотидный состав и определенная последовательность нуклеотидных
звеньев в полимерной цепи.
Например:
В
сокращённом однобуквенном обозначении эта структура записывается как
…– А
– Г – Ц –…
·
Под
вторичной структурой нуклеиновых кислот понимают пространственно
упорядоченные формы полинуклеотидных цепей.
Вторичная
структура ДНК
представляет собой две параллельные неразветвленные полинуклеотидные цепи,
закрученные вокруг общей оси в двойную спираль.
Такая
пространственная структура удерживается множеством водородных связей,
образуемых азотистыми основаниями, направленными внутрь спирали.
Водородные
связи возникают между пуриновым основанием одной цепи и пиримидиновым
основанием другой цепи. Эти основания составляют комплементарные пары (от лат. complementum
— дополнение).
Образование
водородных связей между комплементарными парами оснований обусловлено их пространственным
соответствием.
Пиримидиновое
основание комплементарно пуриновому основанию:
Водородные
связи между другими парами оснований не позволяют им разместиться в структуре
двойной спирали. Таким образом,
·
ТИМИН
(Т) комплементарен АДЕНИНУ (А),
·
ЦИТОЗИН
(Ц) комплементарен ГУАНИНУ (Г).
Комплементарность
оснований определяет комплементарность цепей в молекулах ДНК.
Комплементарность
полинуклеотидных цепей служит химической основой главной функции ДНК – хранения
и передачи наследственных признаков.
Способность
ДНК не только хранить, но и использовать генетическую информацию определяется
следующими ее свойствами:
·
молекулы
ДНК способны к репликации (удвоению), т.е. могут обеспечить возможность синтеза
других молекул ДНК, идентичных исходным, поскольку последовательность оснований
в одной из цепей двойной спирали контролирует их расположение в другой цепи.
·
молекулы
ДНК могут направлять совершенно точным и определенным образом синтез белков,
специфичных для организмов данного вида.
Вторичная
структура РНК
В
отличие от ДНК, молекулы РНК состоят из одной полинуклеотидной цепи и не имеют
строго определенной пространственной формы (вторичная структура РНК зависит от
их биологических функций).
Основная роль РНК – непосредственное участие в биосинтезе белка.
Известны
три вида клеточных РНК, которые отличаются по местоположению в клетке, составу,
размерам и свойствам, определяющим их специфическую роль в образовании белковых
макромолекул:
·
информационные
(матричные) РНК передают закодированную в ДНК информацию о структуре белка от
ядра клетки к рибосомам, где и осуществляется синтез белка;
·
транспортные
РНК собирают аминокислоты в цитоплазме клетки и переносят их в рибосому;
молекулы РНК этого типа «узнают» по соответствующим участкам цепи
информационной РНК, какие аминокислоты должны участвовать в синтезе белка;
·
рибосомные
РНК обеспечивают синтез белка определенного строения, считывая информацию с
информационной (матричной) РНК.
Нуклеиновые кислоты РНК (слева) и ДНК (справа).
Нуклеи́новая кислота (от лат. nucleus — ядро) — высокомолекулярное органическое соединение, биополимер (полинуклеотид), образованный остатками нуклеотидов. Нуклеиновые кислоты ДНК и РНК присутствуют в клетках всех живых организмов и выполняют важнейшие функции по хранению, передаче и реализации наследственной информации.
История исследования[править | править код]
- В 1847 из экстракта мышц быка было выделено[1] вещество, которое получило название «инозиновая кислота». Это соединение стало первым изученным нуклеотидом. В течение последующих десятилетий были установлены детали его химического строения. В частности, было показано, что инозиновая кислота является рибозид-5′-фосфатом, и содержит N-гликозидную связь.
- В 1868 году швейцарским химиком Фридрихом Мишером при изучении некоторых биологических субстанций было открыто неизвестное ранее вещество. Вещество содержало фосфор и не разлагалось под действием протеолитических ферментов. Также оно обладало выраженными кислотными свойствами. Вещество было названо «нуклеином». Соединению была приписана брутто-формула C29H49N9O22P3.
- Уилсон обратил внимание на практическую идентичность химического состава «нуклеина» и открытого незадолго до этого «хроматина» — главного компонента хромосом[2]. Было выдвинуто предположение об особой роли «нуклеина» в передаче наследственной информации.
- В 1889 г Рихард Альтман ввел термин «нуклеиновая кислота», а также разработал удобный способ получения нуклеиновых кислот, не содержащих белковых примесей.
- Левин и Жакоб, изучая продукты щелочного гидролиза нуклеиновых кислот, выделили их основные составляющие — нуклеотиды и нуклеозиды, а также предложили структурные формулы, верно описывающие их химические свойства.
- В 1921 году Левин выдвинул гипотезу «тетрануклеотидной структуры ДНК»[3], оказавшуюся впоследствии ошибочной[4].
- В 1935 году Клейн и Танхаузер с помощью фермента фосфатазы провели мягкое фрагментирование ДНК, в результате чего были получены в кристаллическом состоянии четыре ДНК-образующих нуклеотида[5]. Это открыло новые возможности для установления структуры этих соединений.
- В 1940-е годы научная группа в Кембридже под руководством Александера Тодда проводит широкие синтетические исследования в области химии нуклеотидов и нуклеозидов. В результате их работы были установлены все детали химического строения и стереохимии нуклеотидов. За цикл работ в этой области Александер Тодд был награждён Нобелевской премией в области химии в 1957 году.
- В 1951 году Чаргаффом была установлена закономерность содержания в нуклеиновых кислотах нуклеотидов разных типов, получившая впоследствии название Правило Чаргаффа.
- В 1953 году Уотсоном и Криком установлена вторичная структура ДНК, двойная спираль[6].
Способы выделения[править | править код]
Гелеобразный осадок нуклеиновой кислоты
Описаны многочисленные методики выделения нуклеиновых кислот из природных источников. Основными требованиями, предъявляемыми к методу выделения, являются эффективное отделение нуклеиновых кислот от белков, а также минимальная степень фрагментации полученных препаратов. Классический метод выделения ДНК был описан в 1952 году и используется в настоящее время без значительных изменений[7]. Клеточные стенки исследуемого биологического материала разрушаются одним из стандартных методов, а затем обрабатываются анионным детергентом. При этом белки выпадают в осадок, а нуклеиновые кислоты остаются в водном растворе. ДНК может быть осаждена в виде геля осторожным добавлением этанола к её солевому раствору. Концентрацию полученной нуклеиновой кислоты, а также наличие примесей (белки, фенол) обычно определяют спектрофотометрически по поглощению на А260 нм.
Нуклеиновые кислоты легко деградируют под действием особого класса ферментов — нуклеаз. В связи с этим при их выделении важно обработать лабораторное оборудование и материалы соответствующими ингибиторами. Так, например, при выделении РНК широко используется такой ингибитор рибонуклеаз как DEPC.
Физические свойства[править | править код]
Нуклеиновые кислоты хорошо растворимы в воде, практически нерастворимы в органических растворителях. Очень чувствительны к действию температуры и критическим значениям уровня pH. Молекулы ДНК с высокой молекулярной массой, выделенные из природных источников, способны фрагментироваться под действием механических сил, например, при перемешивании раствора. Нуклеиновые кислоты фрагментируются ферментами — нуклеазами.
Строение[править | править код]
Фрагмент полимерной цепочки ДНК
Полимерные формы нуклеиновых кислот называют полинуклеотидами.
Существуют 4 уровня структурной организации нуклеиновых кислот: первичная, вторичная, третичная и четвертичная. Первичная структура представляет собой цепочки из нуклеотидов, соединяющихся через остаток фосфорной кислоты (фосфодиэфирная связь). Вторичная структура — это две цепи нуклеиновых кислот соединённые водородными связями. Стоит отметить, что цепи соединяются по типу «голова-хвост» (3′ к 5′), по принципу комплементарности (азотистые основания находятся внутри этой структуры). Третичная структура, или же спираль, образуется за счет радикалов азотистых оснований (образуются водородные дополнительные связи, которые и сворачивают эту структуру, тем самым обуславливая её прочность). И наконец 4 структура — это комплексы гистонов и нитей хроматина.
Поскольку в нуклеотидах существует только два типа гетероциклических молекул, рибоза и дезоксирибоза, то и имеется лишь два вида нуклеиновых кислот — дезоксирибонуклеиновая (ДНК) и рибонуклеиновая (РНК).
Мономерные формы также встречаются в клетках и играют важную роль в процессах передачи сигналов или запасании энергии. Наиболее известный мономер РНК — АТФ, аденозинтрифосфорная кислота, важнейший аккумулятор энергии в клетке.
ДНК и РНК[править | править код]
- ДНК (дезоксирибонуклеиновая кислота). Сахар — дезоксирибоза, азотистые основания: пуриновые — гуанин (G), аденин (A), пиримидиновые — тимин (T) и цитозин (C). ДНК часто состоит из двух полинуклеотидных цепей, направленных антипараллельно.
- РНК (рибонуклеиновая кислота). Сахар — рибоза, азотистые основания: пуриновые — гуанин (G), аденин (A), пиримидиновые урацил (U) и цитозин (C). Структура полинуклеотидной цепочки аналогична таковой в ДНК. Из-за особенностей рибозы молекулы РНК часто имеют различные вторичные и третичные структуры, образуя комплементарные участки между разными цепями.
Типы РНК[править | править код]
Ма́тричная рибонуклеи́новая кислота́ (мРНК, синоним — информацио́нная РНК, иРНК) — РНК, содержащая информацию о первичной структуре (аминокислотной последовательности) белков[8]. мРНК синтезируется на основе ДНК в ходе транскрипции, после чего, в свою очередь, используется в ходе трансляции как матрица для синтеза белков. Тем самым мРНК играет важную роль в «проявлении» (экспрессии) генов.
Рибосо́мные рибонуклеи́новые кисло́ты (рРНК) — несколько молекул РНК, составляющих основу рибосомы. Основной функцией рРНК является осуществление процесса трансляции — считывания информации с мРНК при помощи адапторных молекул тРНК и катализ образования пептидных связей между присоединёнными к тРНК аминокислотами.
Структура транспортной РНК
Транспортная РНК, тРНК — рибонуклеиновая кислота, функцией которой является транспортировка аминокислот к месту синтеза белка. Имеет типичную длину от 73 до 93 нуклеотидов и размеры около 5 нм. тРНК также принимают непосредственное участие в наращивании полипептидной цепи, присоединяясь — будучи в комплексе с аминокислотой — к кодону мРНК и обеспечивая необходимую для образования новой пептидной связи конформацию комплекса.
Для каждой аминокислоты существует своя тРНК.
тРНК является одноцепочечной РНК, однако в функциональной форме имеет конформацию «клеверного листа». Аминокислота ковалентно присоединяется к 3′-концу молекулы с помощью специфичного для каждого типа тРНК фермента аминоацил-тРНК-синтетазы.
На участке C находится антикодон, соответствующий аминокислоте.
Некодирующие РНК (non-coding RNA, ncRNA) — это молекулы РНК, которые не транслируются в белки. Ранее использовавшийся синоним, малые РНК (smRNA, small RNA), в настоящее время не используется, так как некоторые некодирующие РНК могут быть очень большими, например, Xist.
Последовательность ДНК, на которой транскрибируются некодирующие РНК, часто называют РНК-геном.
Некодирующие РНК включают в себя молекулы РНК, которые выполняют очень важные функции в клетке — транспортные РНК (тРНК), рибосомные РНК (рРНК), такие малые РНК, как малые ядрышковые РНК (snoRNA), микроРНК, siRNA, piRNA, а также длинные некодирующие РНК — Xist, Evf, Air, CTN, PINK, TUG1.
Последние транскриптомные технологии (секвенирование РНК) и методы ДНК-микрочипов предполагают наличие более 30000 длинных некодирующих РНК (англ. long ncRNA). Примерно такое же количество малых регуляторных РНК содержится в геноме мыши.
Примечания[править | править код]
- ↑ J. Liebig. ??? (неопр.) // Annalen. — 1847. — Т. 62. — С. 257.
- ↑ Edmund B. Wilson. An Atlas of the Fertilization and Karyokinesis of the Ovum. — N. Y.: Macmillan, 1895. — P. 4.
- ↑ P. A. Levene. ??? (англ.) // J. Biol. Chem. : journal. — 1921. — Vol. 48. — P. 119.
- ↑ Во время выдвижения «тетрануклеотидной структуры» химики критически относились к самой возможности существования макромолекул, вследствие чего ДНК была приписана структура с низкой молекулярной массой
- ↑ W. Klein, S. J. Thannhauser. ??? (неопр.) // Z. physiol. Chem.. — 1935. — Т. 231. — С. 96.
- ↑ J. D. Watson, F. H. C. Crick. Molecular Structure of Nucleic Acids: A Structure for Deoxyribose Nucleic Acid (англ.) // Nature. — 1953. — Vol. 171. — P. 737—738. — doi:10.1038/171737a0.
- ↑ Ernest R. M. Kay, Norman S. Simmons, Alexander L. An Improved Preparation of Sodium Desoxyribonucleate (англ.) // J. Am. Chem. Soc. (англ.)русск. : journal. — 1952. — Vol. 74, no. 7. — P. 1724—1726. — doi:10.1021/ja01127a034.
- ↑ Bruce Alberts, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, Peter Walter. Molecular Biology of the Cell. — 5. — Garland Science, 2008. — 1392 с. — ISBN 0815341059.
Литература[править | править код]
- Бартон Д., Оллис У. Д. Общая органическая химия. — М.: Химия, 1986. — Т. 10. — С. 32—215. — 704 с.
- Франк-Каменецкий М. Д. Самая главная молекула. — М.: Наука, 1983. — 160 с.
- Аппель Б., Бенеке И., Бенсон Я., под ред. С. Мюллер. Нуклеиновые кислоты от А до Я. — М.: Бином, 2012. — 352 с. — ISBN 978-5-9963-0376-2.
Азотистые основания и нуклеотиды
Hyклеиновые кислоты играют основную роль в
сохранении и реализации генетической информации (см. с. 234). Различают два типа
нуклеиновых кислот: дезоксирибонуклеиновые кислоты [ДНК (DNA)],
которые обеспечивают сохранение информации, и рибонуклеиновые
кислоты [PHK (RNA)], принимающие участие в процессах генной
экспрессии и биосинтеза белка. Нуклеиновые кислоты построены из
нуклеотидных звеньев, которые в свою очередь состоят из азотистого основания,
углеводного остатка и фосфатной группы. ДНК и РНК различаются по типу
углеводного остатка и структуре оснований.
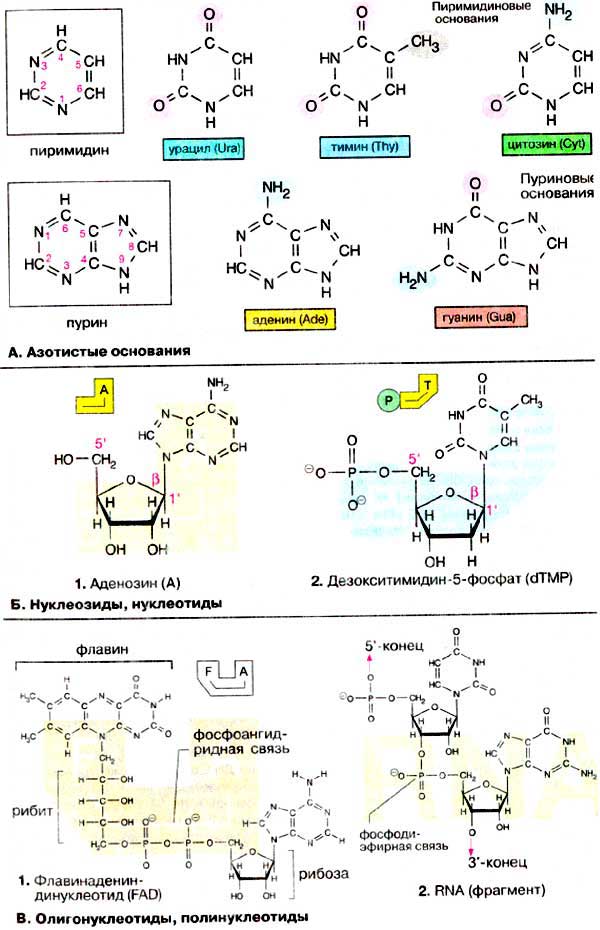
А. Азотистые основания
Азотистые основания — это
ароматические гетероциклические соединения, производные пиримидина
или пурина. Пять соединений этого класса являются основными
структурными компонентами нуклеиновых кислот. Общими для всей живой материи.Пуриновые основания аденин (Ade, но не А) и гуанин
(Guа), а также пиримидиновое основание цитозин (Cyt), входят в состав ДНК и РНК. В состав ДНК входит также тимин (Thy),
5-метил-производное урацила. Основание урацил (Ura) входит только
в состав РНК. В ДНК высших организмов в небольшом количестве присутствует
5-метилцитозин. Производные азотистых оснований присутствуют в тРНК (см.с. 88) и
в других типах РНК.
Б. Нуклеозиды, нуклеотиды
Соединения азотистых оснований с рибозой или 2-дезоксирибозой
(см. с. 44) носят название нуклеозиды. Так, например,
аденин и рибоза образуют нуклеозид аденозин (1,
сокращенно А). Соответствующие производные других азотистых оснований носят
названия гуанозин (G), уридин (U), тимидин (T) и цитидин
(С). Если углеводный остаток представлен 2-дезоксирибозой образуется дезоксинуклеозид,
например 2′-дезоксиаденозин (dA, на схеме не приведен). В клетке 5′-ОН-группа
углеводного остатка нуклеозида этерифицирована фосфорной кислотой. Соответствующее
производное 2′-дезокситимидина (dT), звено ДНК, называется 2′-дезокситимидин-5′-монофосфат
(dTMP) (2). Если 5′-фосфатный остаток
соединяется с другими нуклеозидфосфатными остатками. получаются нуклеозидди-
и нуклеозидтрифосфаты, например АДФ и АТФ — важнейшие коферменты энергообмена
(см. с. 110). Все нуклеозидфосфаты объединяют под общим
названием нуклеотиды.
В нуклеозидах и нуклеотидах пентоза
находится в фуранозной форме (см. с. 40). Углеводный остаток и азотистое
основание связаны N-гликозидной связью между С-1′ углеводного звена и N-9
пуринового или соответственно N-1 пиримидинового цикла. Гликозидная связь
находится в β-конфигурации.
В. Олигонуклеотиды, полинуклеотиды
Остатки фосфорной кислоты могут связываться за счет образования
фосфоангидридной связи. Следовательно, два нуклеотида могут быть связаны через
фосфатные группировки с образованием соответствующего динуклеотида. К
этой группе соединений относятся коферменты [HAДФ+(NADP+)]
и KoA (CoA), а также флавин [ФАД(FAD)] (1,
см. с. 108).
Если фосфатная группа одного нуклеотида взаимодействует с З’-ОН-группой
другого нукпеотида, образуется динуклеотид с фосфодиэфирной связью. Такой
динуклеотид несет на 5′-конце свободную фосфатную группу, а на 3′-конце свободную
ОН-группу. Поэтому можно за счет образования еще одной фосфодиэфирной связи
присоединить новый мононуклеотид. Таким путем образуются олигонуклеотиды
и, наконец, полинуклеотиды.
Полинуклеотиды, составленные из рибонуклеотидных звеньев, называются
рибонуклеиновыми кислотами (РНК), из дезоксирибонуклеотидных мономеров
— дезоксирибонуклеиновыми кислотами (ДНК, см. с. 90).
При обозначении полинуклеотидов указывают сокращенные названия нуклеозидных
звеньев в направлении 5’→3′, т.е. слева направо. Иногда в название
включают фосфатную группу («p»). Так, например, фрагмент РНК, приведенный на
схеме 2, можно записать …pUpG… или сокращенно
..UG…
